

The development of motor coordination in Drosophila embryos
- PMID: 18927150
- PMCID: PMC3501643
- DOI: 10.1242/dev.026773
The development of motor coordination in Drosophila embryos
Abstract
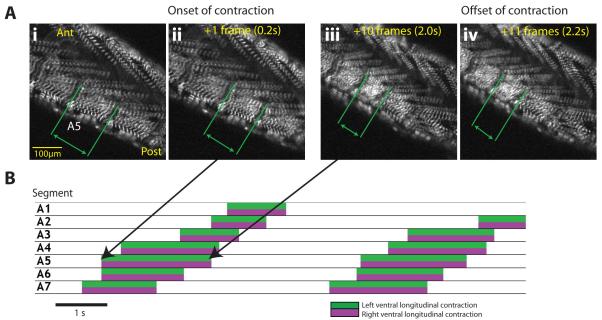
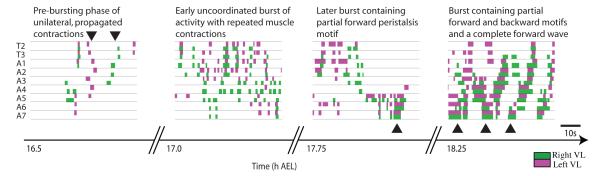
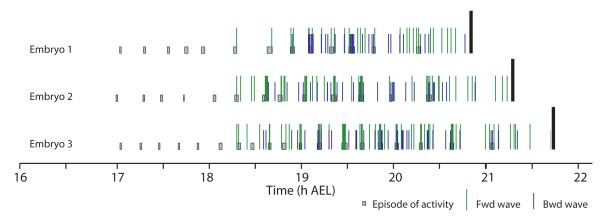
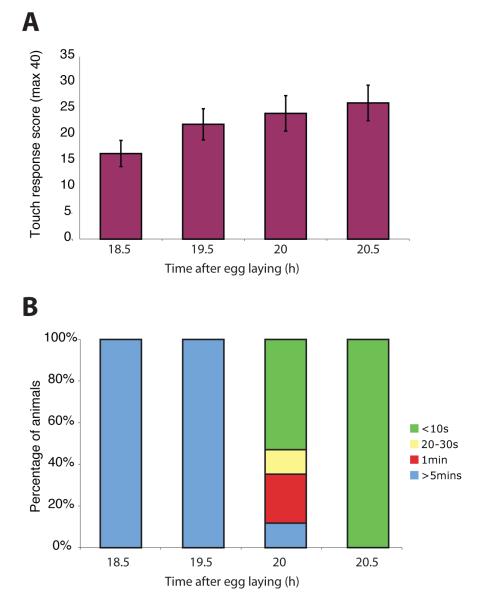
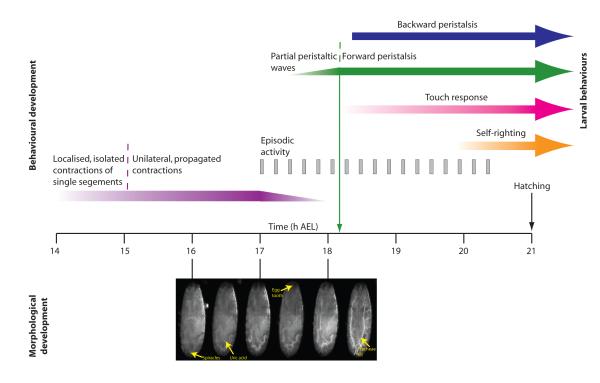
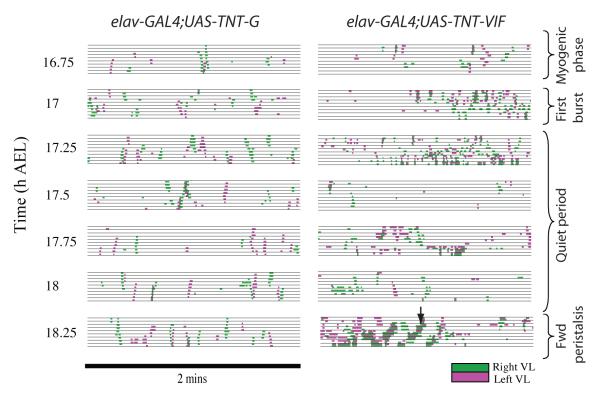
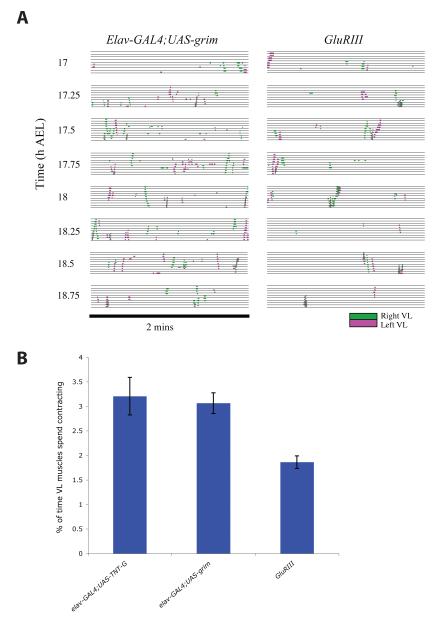
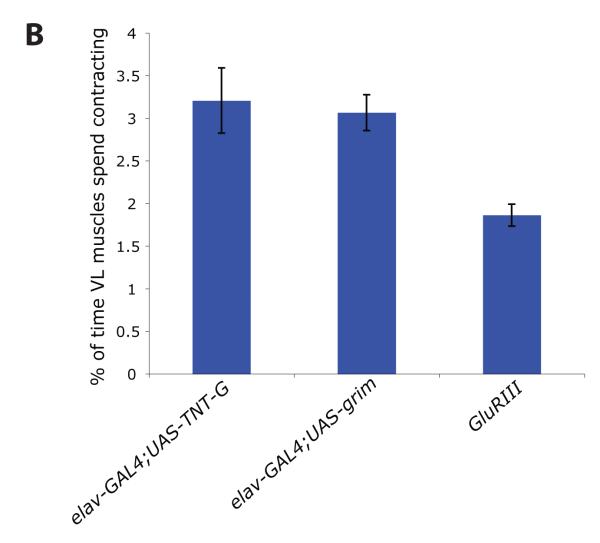
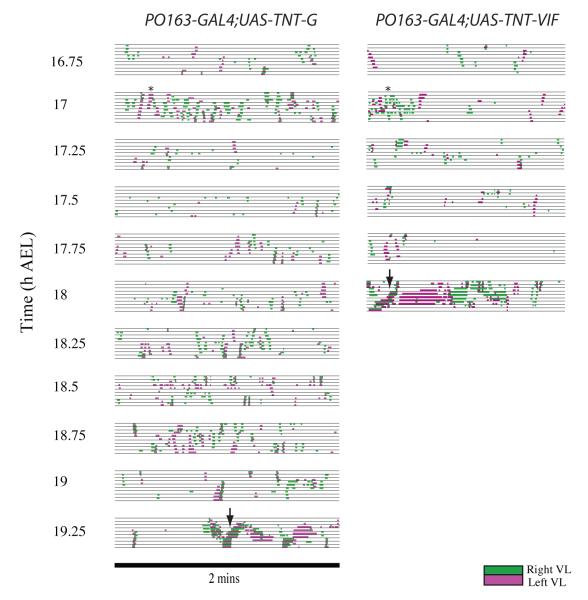
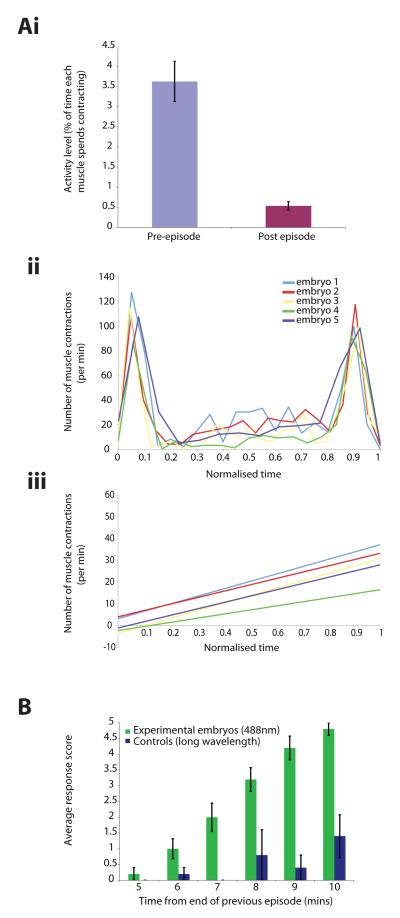
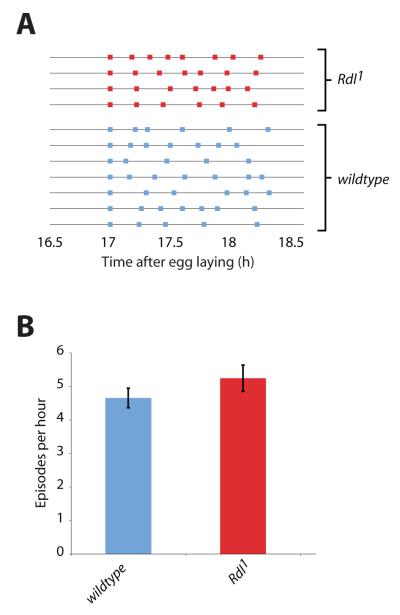
We used non-invasive muscle imaging to study the onset of motor activity and emergence of coordinated movement in Drosophila embryos. Earliest movements are myogenic, and neurally controlled muscle contractions first appear with the onset of bursting activity 17 hours after egg laying. Initial episodes of activity are poorly organised and coordinated crawling sequences only begin to appear after a further hour of bursting. Thus, network performance improves during this first period of activity. The embryo continues to exhibit bursts of crawling-like sequences until shortly before hatching, while other reflexes also mature. Bursting does not begin as a reflex response to sensory input but appears to reflect the onset of spontaneous activity in the motor network. It does not require GABA-mediated transmission, and, by using a light-activated channel to excite the network, we demonstrate activity-dependent depression that may cause burst termination.
Figures
References
-
- Bernhardt RR, Chitnis AB, Lindamer L, Kuwada JY. Identification of spinal neurons in the embryonic and larval zebrafish. J Comp Neurol. 1990;302:603–16. - PubMed
-
- Brustein E, Saint-Amant L, Buss RR, Chong M, McDearmid JR, Drapeau P. Steps during the development of the zebrafish locomotor network. J Physiol Paris. 2003;97:77–86. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
