

Effects of aneuploidy on genome structure, expression, and interphase organization in Arabidopsis thaliana
- PMID: 18927630
- PMCID: PMC2562519
- DOI: 10.1371/journal.pgen.1000226
Effects of aneuploidy on genome structure, expression, and interphase organization in Arabidopsis thaliana
Abstract
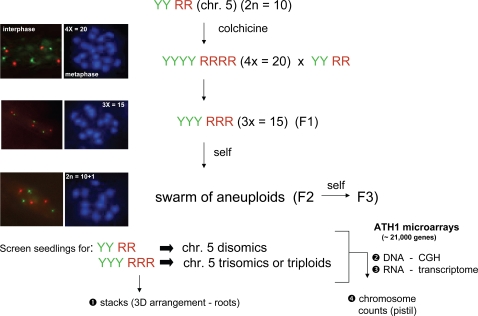
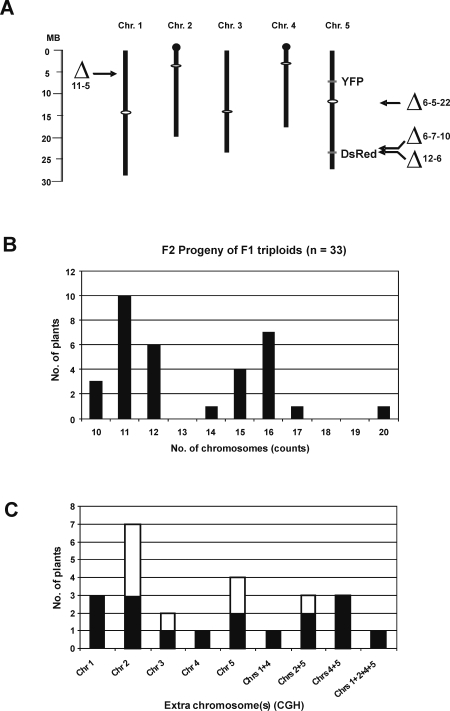
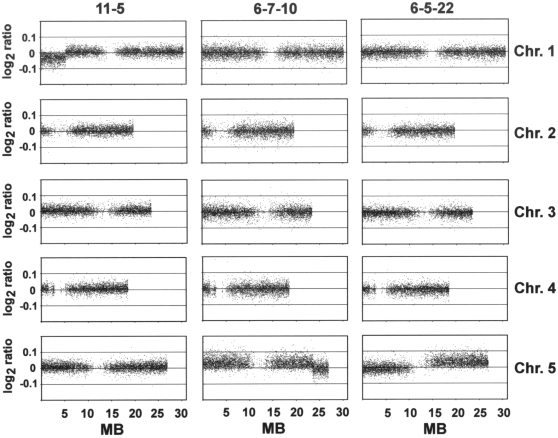
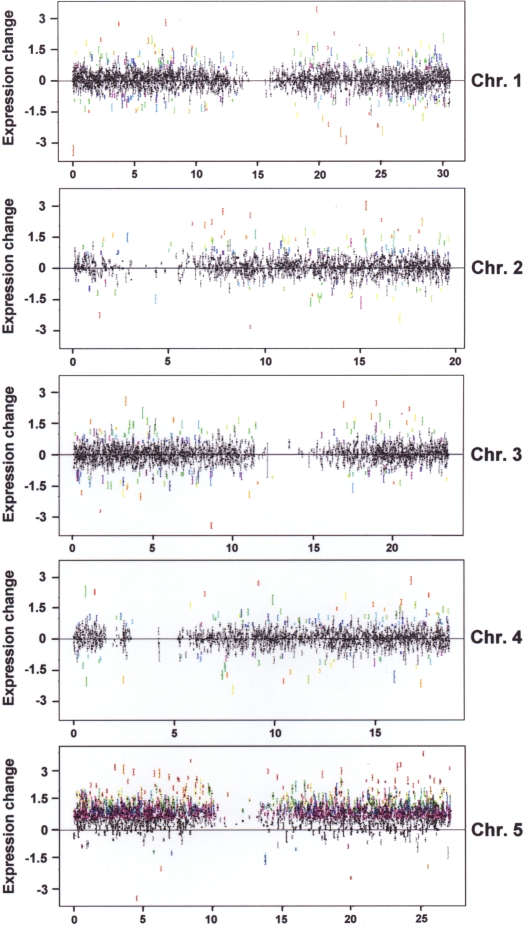
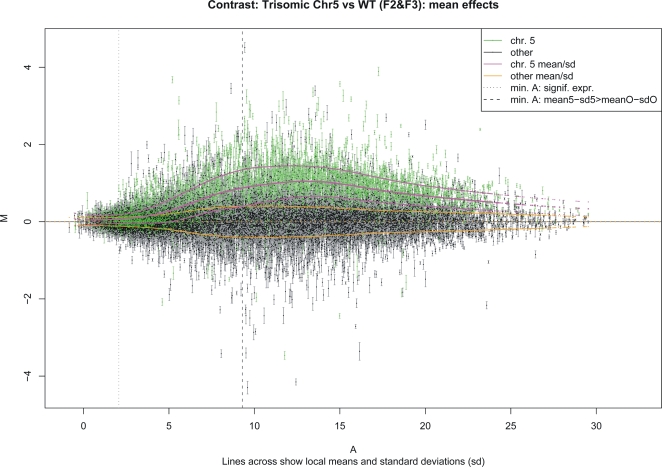
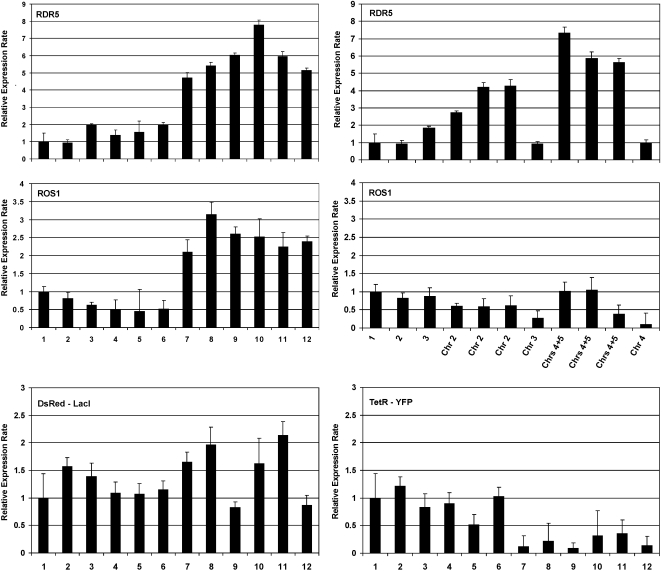
Aneuploidy refers to losses and/or gains of individual chromosomes from the normal chromosome set. The resulting gene dosage imbalance has a noticeable affect on the phenotype, as illustrated by aneuploid syndromes, including Down syndrome in humans, and by human solid tumor cells, which are highly aneuploid. Although the phenotypic manifestations of aneuploidy are usually apparent, information about the underlying alterations in structure, expression, and interphase organization of unbalanced chromosome sets is still sparse. Plants generally tolerate aneuploidy better than animals, and, through colchicine treatment and breeding strategies, it is possible to obtain inbred sibling plants with different numbers of chromosomes. This possibility, combined with the genetic and genomics tools available for Arabidopsis thaliana, provides a powerful means to assess systematically the molecular and cytological consequences of aberrant numbers of specific chromosomes. Here, we report on the generation of Arabidopsis plants in which chromosome 5 is present in triplicate. We compare the global transcript profiles of normal diploids and chromosome 5 trisomics, and assess genome integrity using array comparative genome hybridization. We use live cell imaging to determine the interphase 3D arrangement of transgene-encoded fluorescent tags on chromosome 5 in trisomic and triploid plants. The results indicate that trisomy 5 disrupts gene expression throughout the genome and supports the production and/or retention of truncated copies of chromosome 5. Although trisomy 5 does not grossly distort the interphase arrangement of fluorescent-tagged sites on chromosome 5, it may somewhat enhance associations between transgene alleles. Our analysis reveals the complex genomic changes that can occur in aneuploids and underscores the importance of using multiple experimental approaches to investigate how chromosome numerical changes condition abnormal phenotypes and progressive genome instability.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Duesberg P. Chromosomal chaos and cancer. Sci Am. 2007;296:53–59. - PubMed
-
- Jones PA, Baylin SB. The fundamental role of epigenetic events in cancer. Nat Rev Genet. 2002;3:415–428. - PubMed
-
- Esteller M. Cancer epigenomics: DNA methylomes and histone-modification maps. Nat Rev Genet. 2007;8:286–298. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
