

Ploidy status and copy number aberrations in primary glioblastomas defined by integrated analysis of allelic ratios, signal ratios and loss of heterozygosity using 500K SNP Mapping Arrays
- PMID: 18928532
- PMCID: PMC2576260
- DOI: 10.1186/1471-2164-9-489
Ploidy status and copy number aberrations in primary glioblastomas defined by integrated analysis of allelic ratios, signal ratios and loss of heterozygosity using 500K SNP Mapping Arrays
Abstract
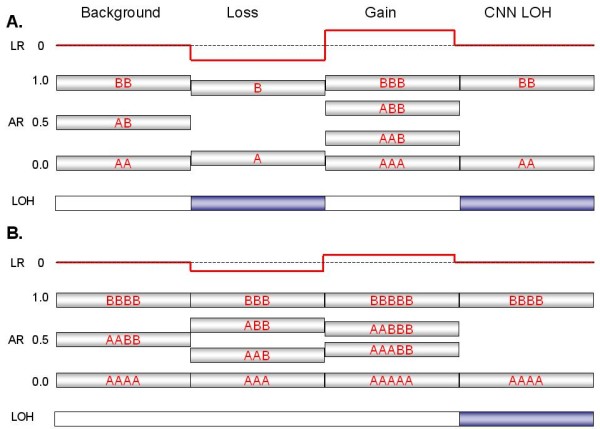
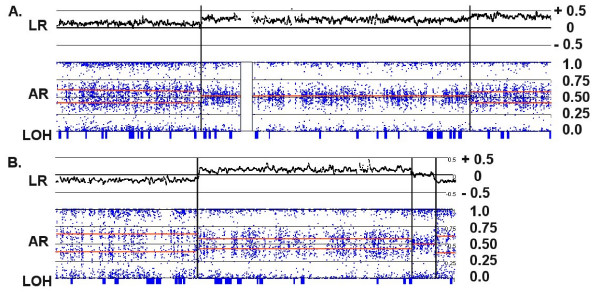
Background: Genomic hybridization platforms, including BAC-CGH and genotyping arrays, have been used to estimate chromosome copy number (CN) in tumor samples by detecting the relative strength of genomic signal. The methods rely on the assumption that the predominant chromosomal background of the samples is diploid, an assumption that is frequently incorrect for tumor samples. In addition to generally greater resolution, an advantage of genotyping arrays over CGH arrays is the ability to detect signals from individual alleles, allowing estimation of loss-of-heterozygosity (LOH) and allelic ratios to enhance the interpretation of copy number alterations. Copy number events associated with LOH potentially have the same genetic consequences as deletions.
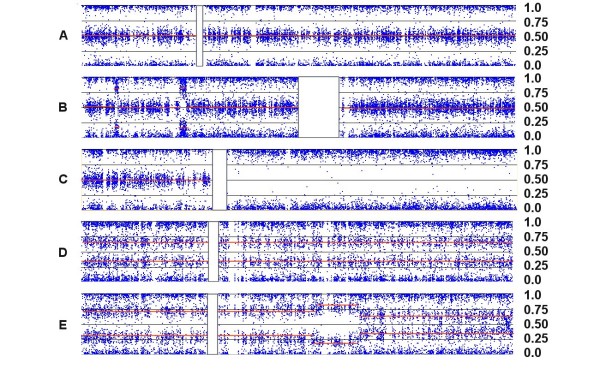
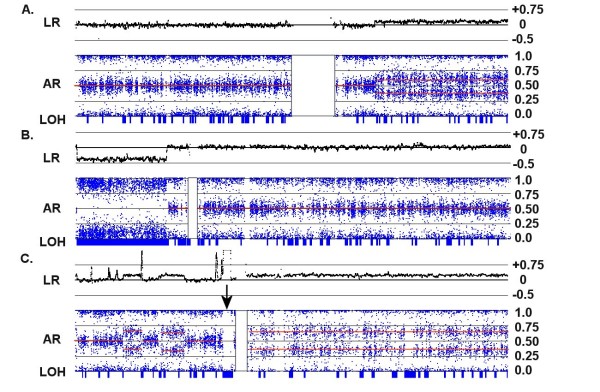
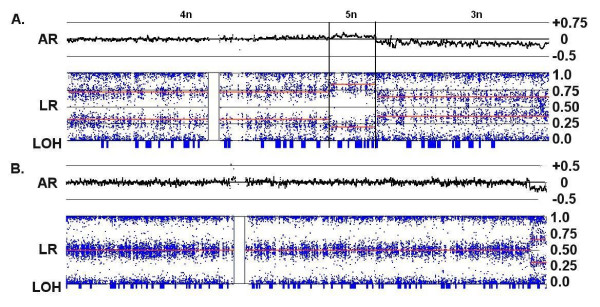
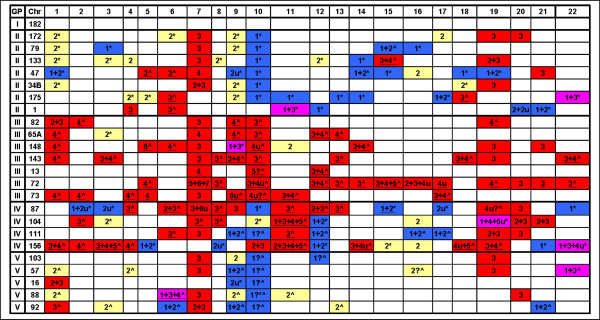
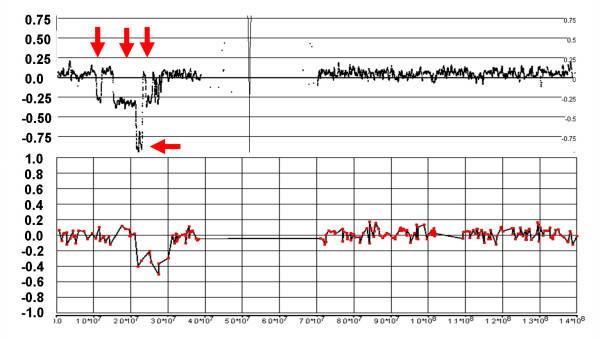
Results: We have utilized allelic ratios to detect patterns that are indicative of higher ploidy levels. An integrated analysis using allelic ratios, total signal and LOH indicates that many or most of the chromosomes from 24 glioblastoma tumors are in fact aneuploid. Some putative whole-chromosome losses actually represent trisomy, and many apparent sub-chromosomal losses are in fact relative losses against a triploid or tetraploid background.
Conclusion: These results suggest a re-interpretation of previous findings based only on total signal ratios. One interesting observation is that many single or multiple-copy deletions occur at common putative tumor suppressor sites subsequent to chromosomal duplication; these losses do not necessarily result in LOH, but nonetheless occur in conspicuous patterns. The 500 K Mapping array was also capable of detecting many sub-mega base losses and gains that were overlooked by CGH-BAC arrays, and was superior to CGH-BAC arrays in resolving regions of complex CN variation.
Figures
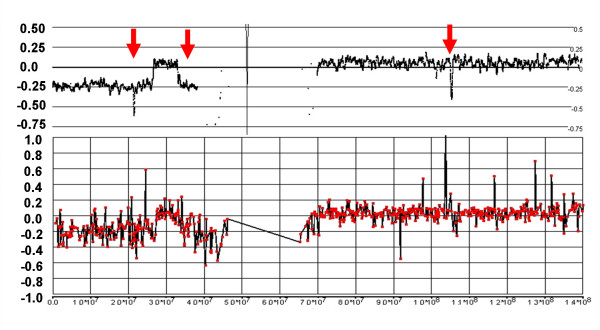
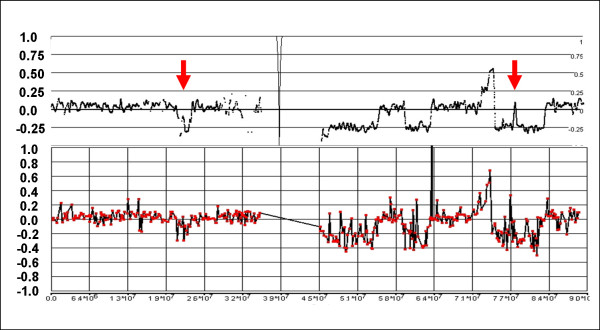
References
-
- Cavenee WK, Dryja TP, Phillips RA, Benedict WF, Godbout R, Gallie BL, Murphree AL, Strong LC, White RL. Expression of recessive alleles by chromosomal mechanisms in retinoblastoma. Nature. 1983;305:779–784. - PubMed
-
- Lo KC, Rossi MR, LaDuca J, Hicks DG, Turpaz Y, Hawthorn L, Cowell JK. Candidate glioblastoma development gene identification using concordance between copy number abnormalities and gene expression level changes. Genes Chromosomes Cancer. 2007;46:875–894. - PubMed
-
- Lo KC, Rossi MR, Burkhardt T, Pomeroy SL, Cowell JK. Overlay analysis of the oligonucleotide array gene expression profiles and copy number abnormalities as determined by array comparative genomic hybridization in medulloblastomas. Genes Chromosomes Cancer. 2007;46:53–66. - PubMed
-
- Cowell JK, Nowak NJ. High-resolution analysis of genetic events in cancer cells using bacterial artificial chromosome arrays and comparative genome hybridization. Adv Cancer Res. 2003;90:91–125. - PubMed
-
- Pinkel D, Albertson DG. Comparative genomic hybridization. Annu Rev Genomics Hum Genet. 2005;6:331–354. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
