

Review
doi: 10.1016/S0065-2911(08)00001-5.
Complex regulatory pathways coordinate cell-cycle progression and development in Caulobacter crescentus
Affiliations
- PMID: 18929067
- PMCID: PMC2621326
- DOI: 10.1016/S0065-2911(08)00001-5
Item in Clipboard
Review
Complex regulatory pathways coordinate cell-cycle progression and development in Caulobacter crescentus
Adv Microb Physiol.
2009.
Abstract
Caulobacter crescentus has become the predominant bacterial model system to study the regulation of cell-cycle progression. Stage-specific processes such as chromosome replication and segregation, and cell division are coordinated with the development of four polar structures: the flagellum, pili, stalk, and holdfast. The production, activation, localization, and proteolysis of specific regulatory proteins at precise times during the cell cycle culminate in the ability of the cell to produce two physiologically distinct daughter cells. We examine the recent advances that have enhanced our understanding of the mechanisms of temporal and spatial regulation that occur during cell-cycle progression.
Figures
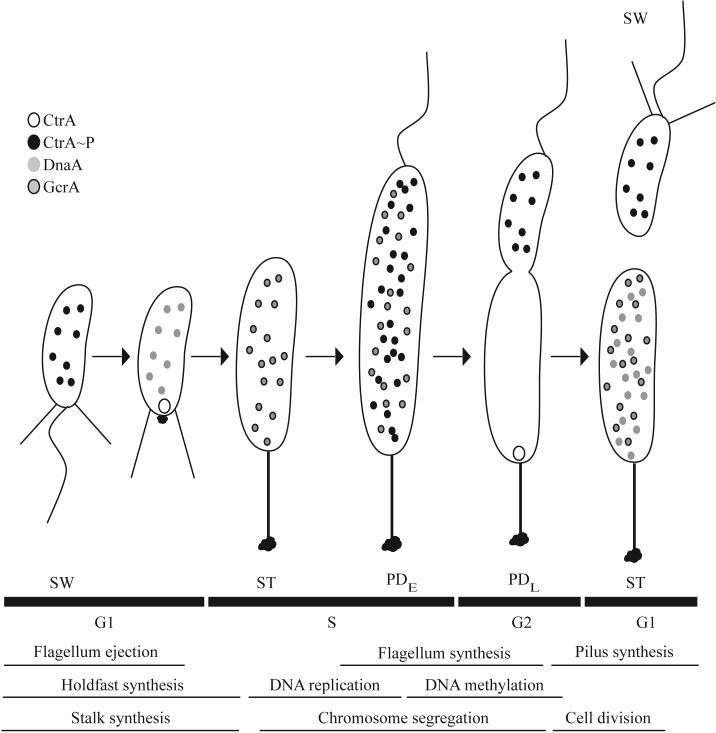
The C. crescentus cell cycle and the localization of the major regulators CtrA, DnaA, and GcrA. The cell cycle, including the swarmer (SW), stalked (ST), early predivisional (PDE), late predivisional (PDL), and progeny cells, is depicted. The stages of the cell division cycle and cell cycle events are shown below the cell cycle schematic. G1 is the pre-synthesis gap period, S is the DNA synthesis period, and G2 is the post-synthesis gap period. Phosphorylated CtrA (CtrA∼P) is found in the swarmer cell. As the cell undergoes the swarmer to stalked cell differentiation, CtrA is recruited to the flagellar pole where it is proteolyzed. DnaA is then synthesized and leads to the production of GcrA, the dominant regulator in the stalked cell. In the predivisional cell, GcrA is responsible for the activation of ctrA transcription. CtrA is phosphorylated and represses transcription of gcrA. In the stalked compartment, CtrA is subject to proteolysis at the stalked pole and both GcrA and DnaA accumulate in the new stalked cell. In the swarmer compartment, phosphorylated CtrA is present and blocks chromosome replication in the new swarmer cell.
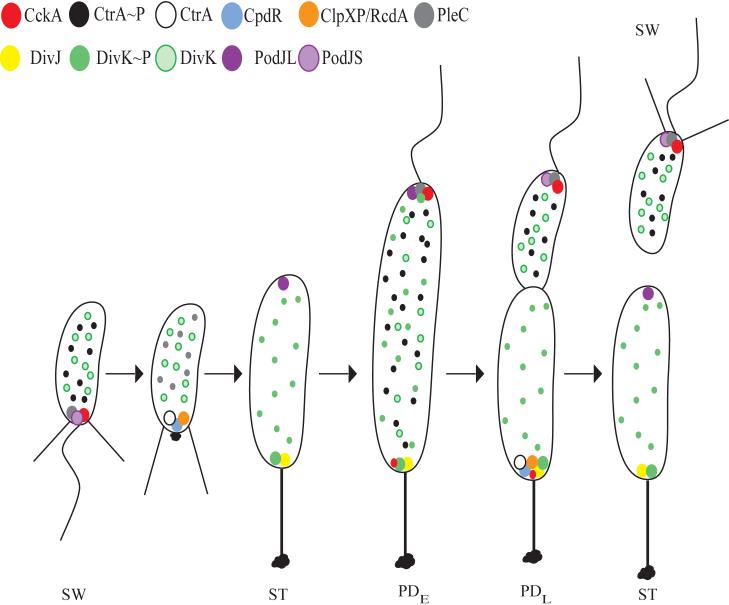
Localization of proteins affecting CtrA phosphorylation and proteolysis during the C. crescentus cell cycle. All stages of the cell cycle, including the swarmer (SW), stalked (ST), early predivisional (PDE), late predivisional (PDL), and progeny cells, are shown. In the swarmer cell, PodJS is localized at the flagellar pole and is responsible for the localization of PleC. PleC dephosphorylates DivK∼P leading to the accumulation of delocalized DivK. CckA is also localized to the flagellar pole and controls CtrA prosphorylation (CtrA∼P), leading to a block in chromosome replication. As the cell undergoes the swarmer to stalked cell differentiation, PodJ is degraded, releasing PleC from the pole. CpdR is localized to the flagellated pole and recruits both CtrA and ClpXP/RcdA to the pole, resulting in the proteolysis of CtrA. In the stalked cell, PodJL is synthesized and localizes to the incipient swarmer pole. DivJ is localized to the stalked pole where it recruits and phosphorylates DivK, leading to accumulation of DivK∼P at the pole and in the cytoplasm. In the early predivisional cell, PodJL recruits PleC to the incipient swarmer pole where PleC dephosphorylates DivK∼P. Meanwhile, DivJ is localized to the stalked pole where DivK is phosphorylated. Due to the opposing actions of PleC and DivJ, DivK and DivK∼P are found in the cytoplasm. In the swarmer compartment of the late predivisional cell, the periplasmic protease PerP is responsible for the cleavage of PodJL to PodJS at the incipient flagellar pole. CckA and PleC remain at the flagellar pole and CtrA is phosphorylated. The absence of DivJ and the presence of PleC in the swarmer compartment lead to the accumulation of unphosphorylated DivK. In the stalked compartment of the predivisional cell, CpdR and ClpXP/RcdA join DivJ and DivK∼P at the stalked pole, resulting in the degradation of CtrA and accumulation of DivK∼P. In the new stalked cell, PodJL is synthesized, CtrA is completely degraded, and CpdR and ClpXP/RcdA are no longer polarly localized.
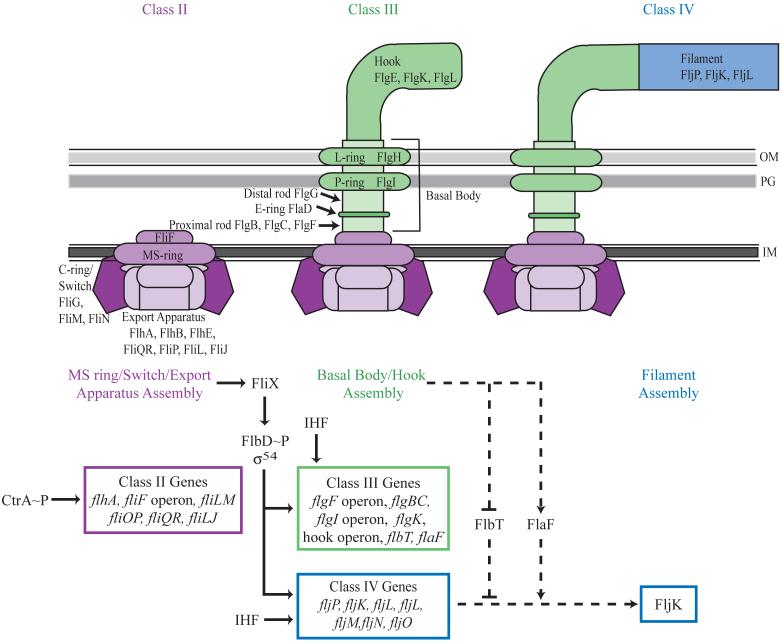
Transcriptional regulation, translational regulation, and assembly of flagellar proteins. The presence of phosphorylated CtrA (CtrA∼P) leads to the transcription of the class II flagellar genes, which include flbD, which is part of the fliF operon. FlbD, aσ54-dependent transcriptional activator, is repressed by FliX until assembly of the class II flagellar proteins, including the export apparatus, C-ring/switch, and MS-ring, is complete. The assembly of class II flagellar proteins is detected by FliX through an unknown mechanism and leads to the activation of FlbD by phosphorylation in a FliX dependent manner. FlbD phosphorylation results in the transcriptional activation of the class III and IV flagellar genes. Maximal transcription of class III and IV flagellar genes also requires IHF. The class III flagellar proteins are then assembled to form the basal body and hook of the flagellum. Although the class IV genes are transcribed, they are subject to post-transcriptional regulation by FlbT and FlaF, which prevent translation of at least one class IV flagellin message, fljK, until assembly of the basal body and hook is complete. Following assembly of the basal body and hook, fljK transcripts are stabilized by FlaF. The presence of FljK, the major flagellin required for motility, enables filament assembly. With the addition of the filament, flagellum assembly is complete. Solid lines depict pathways involved in transcriptional control and dashed lines indicate pathways for translational control. IM, inner membrane; PG, peptidoglycan; OM, outer membrane.
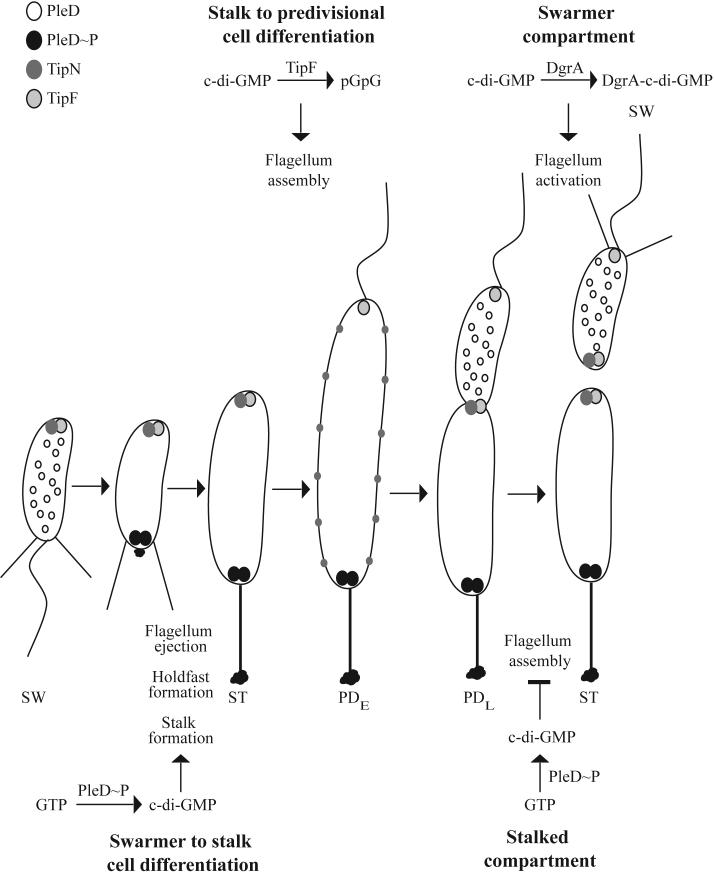
Cyclic-di-GMP signaling throughout the C. crescentus cell cycle. Each stage of the cell cycle, including the swarmer (SW), stalked (ST), early predivisional (PDE), late predivisional (PDL), and progeny cells, is shown. In the swarmer cell PleD is delocalized. As the cell undergoes the swarmer to stalked cell differentiation, dimers of PleD∼P are localized to the flagellar pole leading to the production of cyclic-di-GMP. The increase in c-di-GMP levels promotes flagellum ejection, holdfast formation, and stalk formation. Flagellum assembly in the early predivisional cell is achieved by a reduction in c-di-GMP levels through degradation into linear diguanylate (pGpG) by TipF and occurs at the pole marked by the presence of TipN. After flagellum biosynthesis, TipN is briefly delocalized. In the late predivisional cell, just prior to cell division, flagellum activation occurs as c-di-GMP levels decrease due to binding by proteins such as DgrA. TipF is localized to the pole opposite the flagellum throughout the cell cycle, until the onset of cell division, when both TipN and TipF localize to the mid-cell and mark the new pole following cell separation.
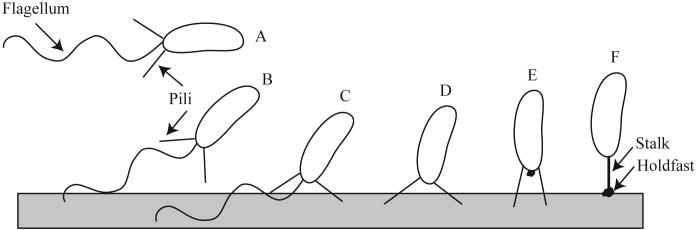
Attachment of a C. crescentus cell to a surface. The stages of initial attachment are shown, A-F. The surface for attachment is shown as a grey box and the polar structures are labeled. Contact with the surface is mediated by flagellar motility, which overcomes repulsive forces (A-B). The flagellum enhances the initial attachment of C. crescentus to the surface (C). Following flagellum ejection, the interaction of the pili with the surface may properly position the cell prior to a more permanent attachment (D). As the swarmer cell differentiates into a stalk cell, the holdfast is exported and the pili are hypothesized to retract (E). The biosynthesis of the stalk brings the holdfast in contact with the surface (F). The holdfast is responsible for the remarkably strong adhesion of C. crescentus to surfaces.
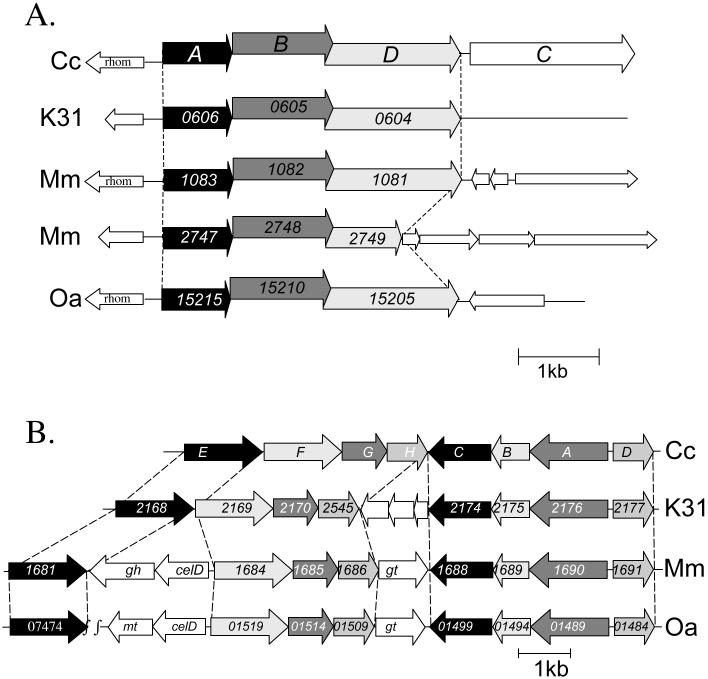
Organization of the hfa and hfs loci in C. crescentus, Caulobacter sp. K31, M. maris and O. alexandrii. Each arrow represents a gene and the direction of transcription for that gene. Cc, C. crescentus CB15; K31, Caulobacter sp. K31; Mm, M. maris MCS10; Oa, Oceanicaulis alexandrii HTCC2633 (A) Comparison of the holdfast attachment loci for C. crescentus, M. maris and O. alexandrii. The holdfast attachment locus is composed of hfaA, hfaB, hfaD and hfaC. The corresponding homologs for hfaA (black), hfaB (dark grey) and hfaD (light grey) in Caulobacter sp. K31, M. maris and O. alexandrii are shown below the hfa locus for C. crescentus. The gene numbers are indicated in the arrow representing the gene for Caulobacter sp. K31, M. maris and O. alexandrii. Predicted open reading frames surrounding the hfa loci are represented with white arrows. There is a conserved rhomboid protease (rhom) upstream of many of the hfa loci. M. maris has two hfa loci. HfaC is not present in any of the other hfa loci. (B) Comparison of the holdfast biosynthesis loci from C. crescentus, Caulobacter sp. K31, M. maris and O. alexandrii. The holdfast biosynthesis and transport loci are comprised of hfsE, hfsF, hfsG, hfsH and then hfsD, hfsA, hfsB and hfsC. The gene numbers for each gene are indicated in the arrow representing the gene for Caulobacter sp. K31, M. maris and O. alexandrii. Both M. maris and O. alexandrii have additional genes that may function in holdfast biosynthesis, which are shown as white arrows. Caulobacter sp. K31 has additional genes within the holdfast biosynthesis and secretion loci (shown in white), but their suggested functions do not appear to be related to polysaccharide biosynthesis. gt indicates a predicted glycosyl transferase; mt, is a predicted mannosyl transferase; gh, a predicted glycosyl hydrolase; celD indicates a gene involved in cellulose biosynthesis. The gene function assignments are based on the annotations of TIGR or JGI and blastp analysis. O. alexandrii has a homolog to hfsE that is located elsewhere in the genome as indicated by the gene number.
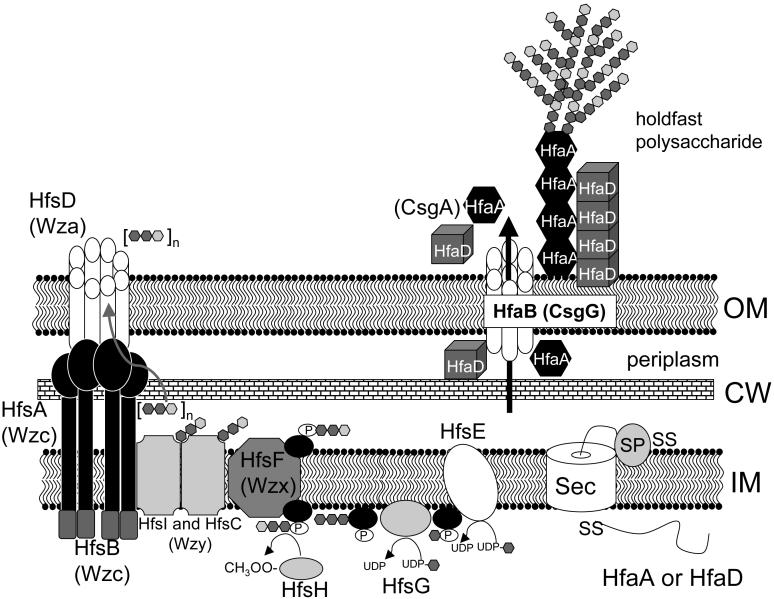
Model of holdfast biosynthesis and attachment in C. crescentus. HfsE transfers N-acetylglucosamine (NAG) from UDP-NAG to undecaprenol (black oval). HfsG transfers NAG to the first sugar on the undecaprenol. HfsH deacetylates some of the NAG on the growing polysaccharide chain to glucosamine. HfsF is a flippase and transfers the small saccharide repeat unit linked to undecaprenol across the cytoplasmic membrane. HfsC and HfsI are polymerases that link the saccharide repeat units together to create the holdfast polysaccharide. HfsA, HfsB and HfsD translocate the holdfast polysaccharide across the outer membrane so it can be anchored to the cell surface via the holdfast attachment proteins. An HfsA tetramer and HfsD octomer create the translocator and HfsB is an ATPase important for phosphorylation and signaling. For the holdfast attachment proteins, HfaA and HfaD are translocated via the Sec system across the cytoplasmic membrane to the periplasm where the signal sequence is cleaved by signal peptidase (SP). After processing, HfaA and HfaD interact with HfaB, the outer membrane secretin. HfaB translocates HfaA and HfaB across the outer membrane where they create the holdfast anchor and associate with the holdfast polysaccharide (only drawn for HfaA) by an unknown mechanism. Homologs to each of the Hfs and Hfa proteins are shown in parentheses adjacent to each protein. UDP, uridine diphosphate; P, phosphate; dark gray hexagon, NAG; light gray hexagon, glucosamine; IM, inner membrane; OM, outer membrane; CW, cell wall; SS, signal sequence.
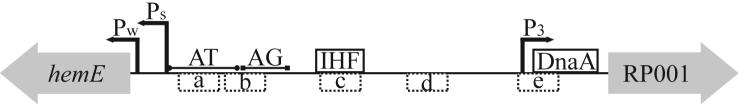
The C. crescentus origin of replication. The region required for chromosome replication is shown as a thick black line and is flanked by the hemE and RP001 genes which are depicted as grey arrows that indicate polarity. The black arrows indicate RNA start sites for the transcription promoters, PW, PS, and P3. The AT and purine (AG) rich regions required for chromosome replication are shown as labeled thin solid bars. The binding sites of IHF and DnaA are shown in solid boxes. The CtrA binding sites are shown in dashed boxes and are labeled with the site designation, a-e.
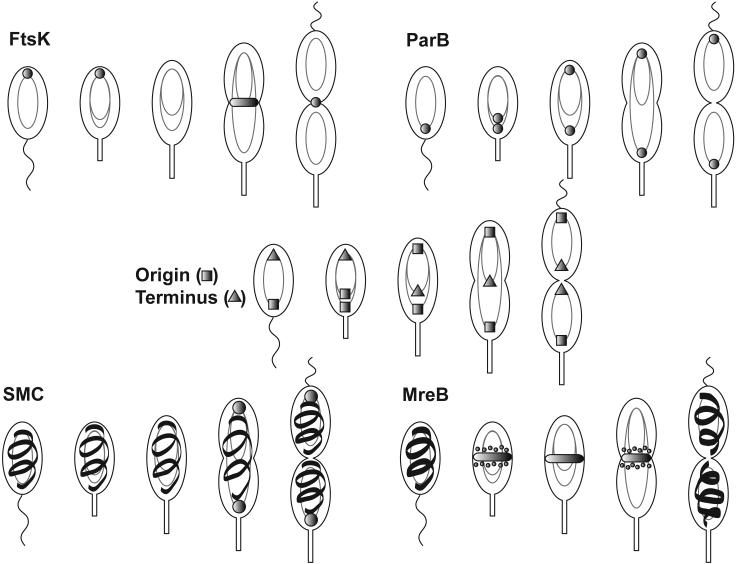
Localization of replication origin and terminus and chromosome segregation proteins in C. crescentus. In each cell cycle, the light gray oval represents a non-replicating chromosome and the tetha structures represent a replicating chromosome. The origin of replication and the terminus (square and triangle in the middle cell cycle, respectively) are at opposite poles in swarmer cells, with the origin at the flagellated pole. A copy of the origin is rapidly moved to the opposite pole early in chromosome replication. The terminus progressively moves to the midcell and the two copies of the terminus are decatenated prior to cell division. The various proteins involved in chromosome segregation are represented by shaded shapes. FtsK localizes to the pole opposite the flagellum and the stalk in swarmer and stalked cells. During constriction, FtsK localizes to the mid-cell where it remains until the cell divides to stabilize the Z-ring and help complete chromosome segregation. ParB co-localizes with and tracks the origin of replication. The SMC protein has a random or spiral-like localization localization throughout the cytoplasm; however during cell division the SMC proteins additionally form discreet foci at the poles. MreB forms a spiral in swarmer cells that coalesces into a band at the mid-cell during division. MreB then returns to a spiral near the end of cell division.
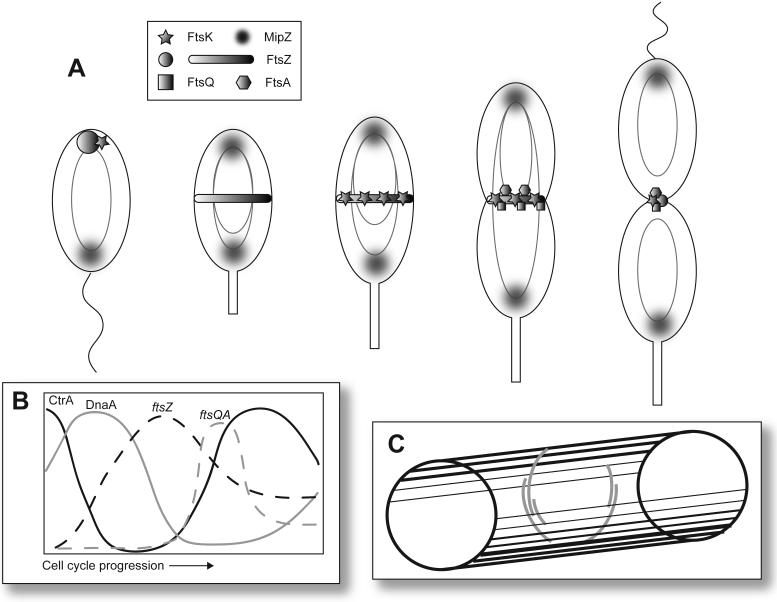
Localization and regulation of cell division proteins in C. crescentus: (A) In each cell, the light gray oval represents a non-replicating chromosome and the tetha structures represent a replicating chromosome. FtsZ (circle) initially localizes to the pole opposite the flagellum along with FtsK (star) in the swarmer cell. MipZ (diffuse shaded circle) resides at the flagellar pole. As the cell cycle is initiated, MipZ tracks with the newly replicated origins and rapidly localizes to both poles. Arrival of MipZ to the pole opposite the stalk displaces FtsZ, which migrates to localize at the midcell (Z-ring, shaded band at the mid-cell). As the cell elongates, FtsA and FtsQ initially localize in a spiral-like pattern (not shown) and FtsK localizes to the midcell. In late predivisional cells, a preponderance of FtsQ (square) and FtsA (hexagon) join the Z-ring. (B) Timing of transcription of ftsZ, ftsQ, and ftsA and variation in abundance of the transcriptional regulators CtrA and DnaA during the cell cycle. The solid lines indicate the proteins levels of the global regulators CtrA and DnaA. The dashed lines represent the transcription pattern of ftsZ and ftsQA as the cell proceeds through the cell cycle. (C) The diagram depicts how multiple short filaments of FtsZ form the Z-ring at the mid-cell. The cell is represented by the cylinder and the FtsZ filaments are represented by the gray curved lines.
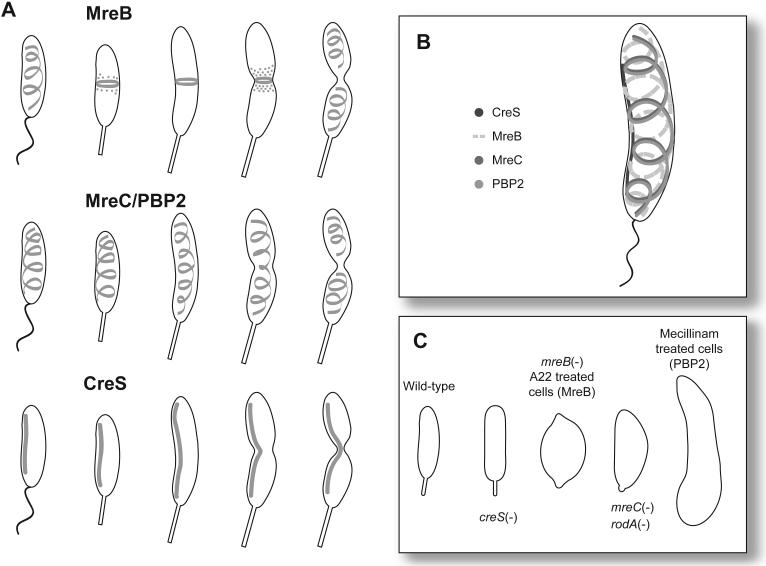
Cell shape proteins in C. crescentus. (A) Localization of cell shape proteins during the cell cycle exhibits. The proteins are represented in gray. MreB localizes as a spiral in swarmer cells, coalesces to a ring in the stalked and predivisional cells, and returns to a spiral at the end of cell division. MreC and PBP2 localize in a similar spiral-like pattern throughout the cell cycle. CreS aligns along the inner curve of the cell throughout the cell cycle. (B) Cell shape protein localization in swarmer cells. MreC and PBP2 co-align while the MreB spiral is offset from the MreC/PBP2 spirals. CreS lies along the inner curve of the bacterium. (C) Cell shapes resulting from the absence or inhibition of cytoskeletal proteins. Gene deletions and protein depletions are indicated by (-). A22 targets MreB and cells grown in its presence have an identical phenotype to cells depleted for MreB. PBP2 is inhibited by the addition of mecillinam.
References
-
- Aaron M, Charbon G, Lam H, Schwarz H, Vollmer W, Jacobs-Wagner C. The tubulin homologue FtsZ contributes to cell elongation by guiding cell wall precursor synthesis in Caulobacter crescentus. Mol. Microbiol. 2007;64:938–952. - PubMed
-
- Abraham WR, Strompl C, Meyer H, Lindholst S, Moore ER, Christ R, Vancanneyt M, Tindall BJ, Bennasar A, Smit J, Tesar M. Phylogeny and polyphasic taxonomy of Caulobacter species. Proposal of Maricaulis gen. nov. with Maricaulis maris (Poindexter) comb. nov. as the type species, and emended description of the genera Brevundimonas and Caulobacter. Int. J. Syst. Bacteriol. 1999;49(Pt 3):1053–73. - PubMed
-
- Addinall SG, Holland B. The tubulin ancester, FtsZ, draughtsman, designer and driving force for bacterial cytokinesis. J. Mol. Biol. 2002;318:219–236. - PubMed
-
- Aldridge P, Hughes KT. Regulation of flagellar assembly. Currents Opinion in Microbiology. 2002;5:160–5. - PubMed
-
- Aldridge P, Jenal U. Cell cycle-dependent degradation of a flagellar motor component requires a novel-type response regulator. Mol. Microbiol. 1999;32:379–392. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
