

Methods for studying store-operated calcium entry
- PMID: 18929662
- PMCID: PMC2643845
- DOI: 10.1016/j.ymeth.2008.09.009
Methods for studying store-operated calcium entry
Abstract
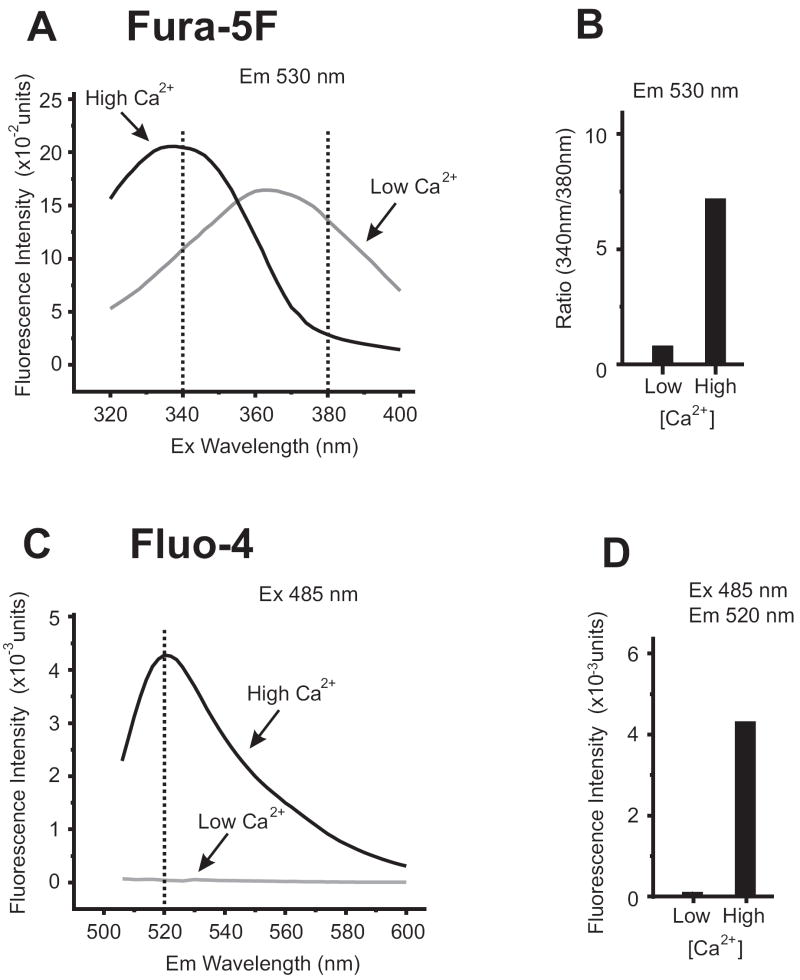
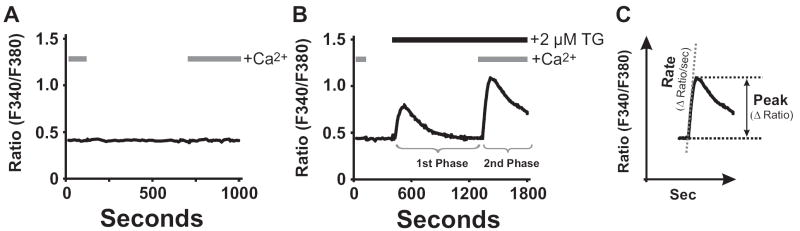
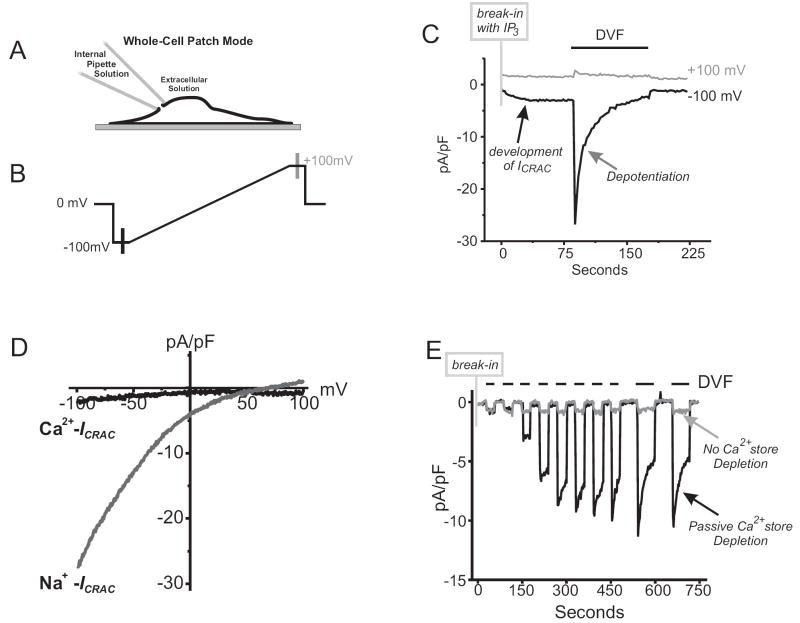
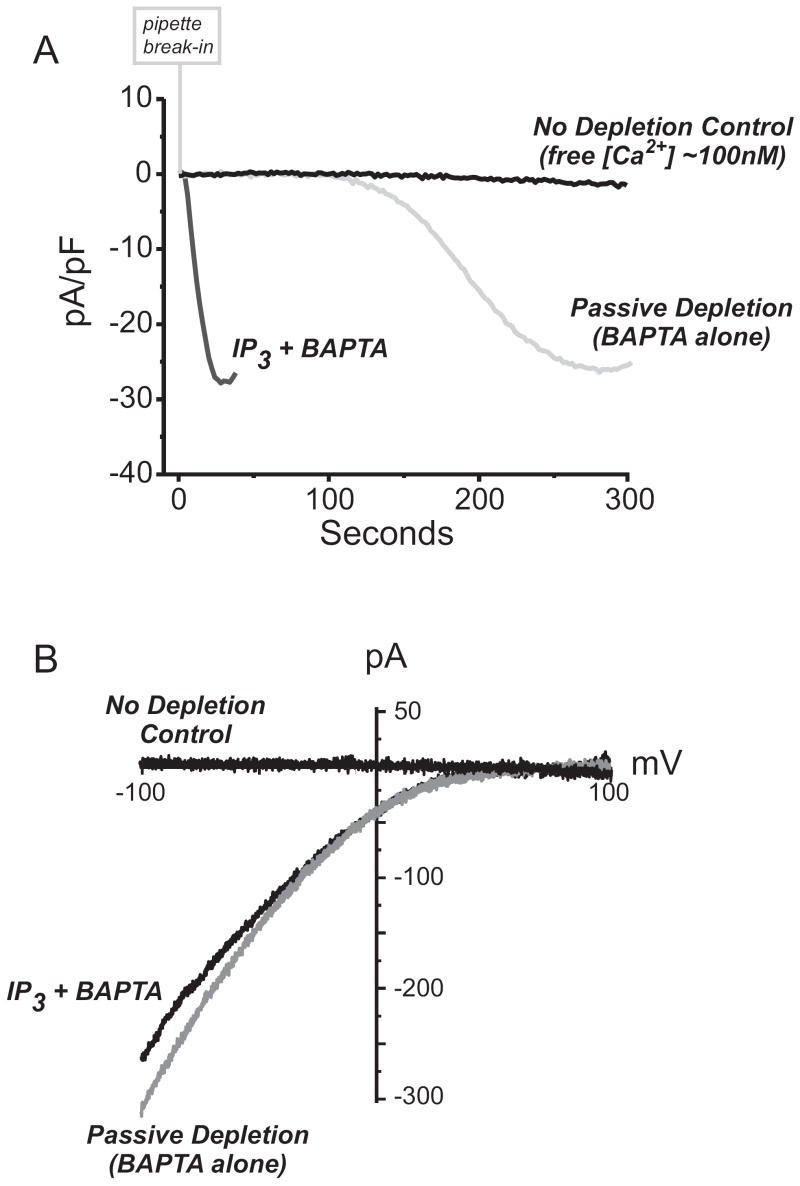
Activation of surface membrane receptors coupled to phospholipase C results in the generation of cytoplasmic Ca2+ signals comprised of both intracellular Ca2+ release, and enhanced entry of Ca2+ across the plasma membrane. A primary mechanism for this Ca2+ entry process is attributed to store-operated Ca2+ entry, a process that is activated by depletion of Ca2+ ions from an intracellular store by inositol 1,4,5-trisphosphate. Our understanding of the mechanisms underlying both Ca2+ release and store-operated Ca2+ entry have evolved from experimental approaches that include the use of fluorescent Ca2+ indicators and electrophysiological techniques. Pharmacological manipulation of this Ca2+ signaling process has been somewhat limited; but recent identification of key molecular players, STIM and Orai family proteins, has provided new approaches. Here we describe practical methods involving fluorescent Ca2+ indicators and electrophysiological approaches for dissecting the observed intracellular Ca2+ signal to reveal characteristics of store-operated Ca2+ entry, highlighting the advantages, and limitations, of these approaches.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
