

Intergenerational and striatal CAG repeat instability in Huntington's disease knock-in mice involve different DNA repair genes
- PMID: 18930147
- PMCID: PMC2811282
- DOI: 10.1016/j.nbd.2008.09.014
Intergenerational and striatal CAG repeat instability in Huntington's disease knock-in mice involve different DNA repair genes
Abstract
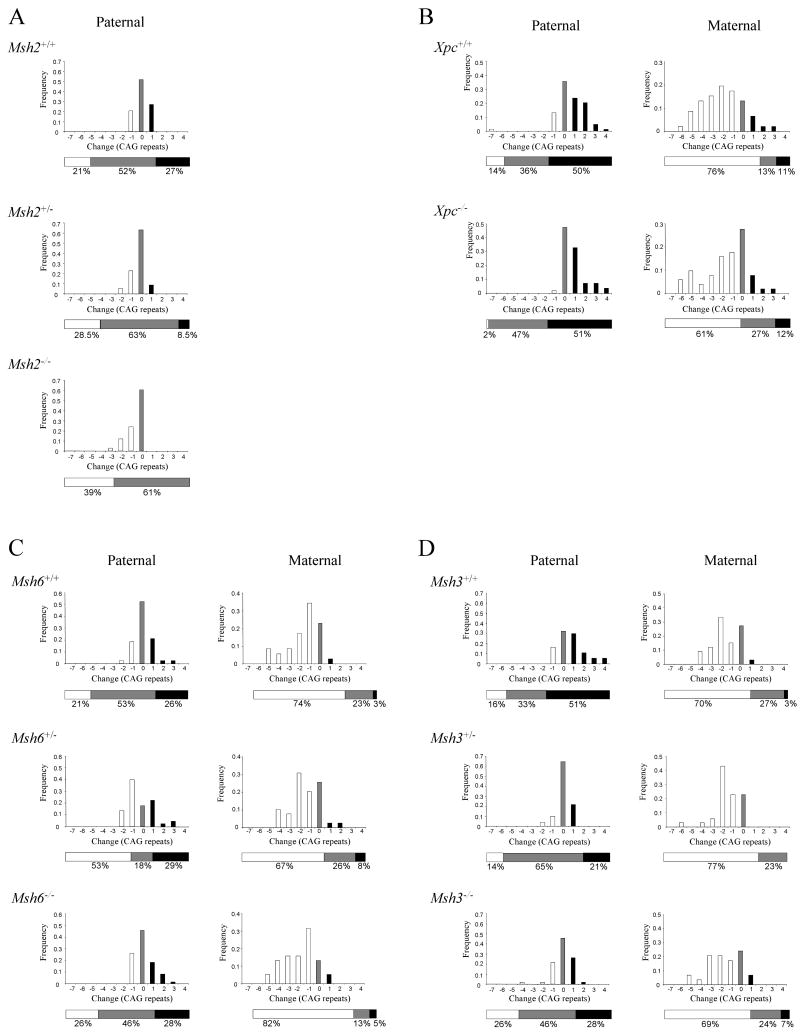
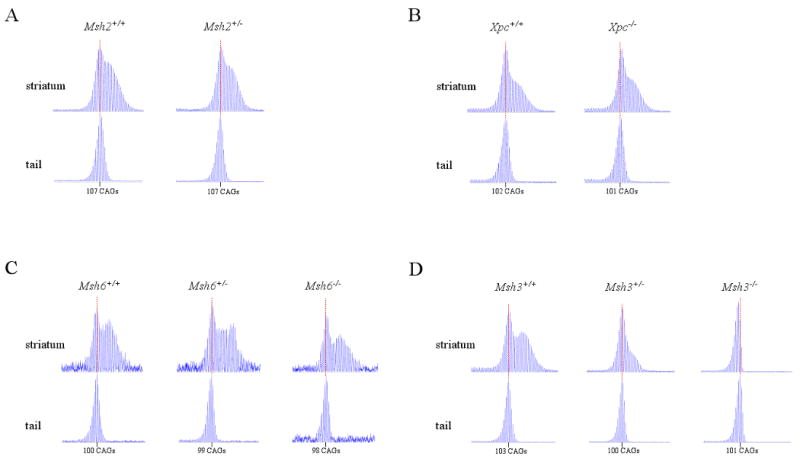
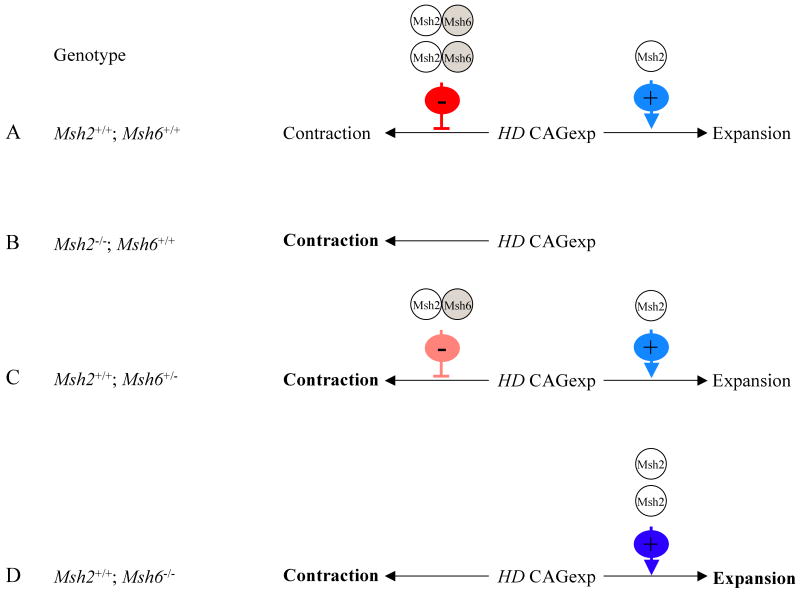
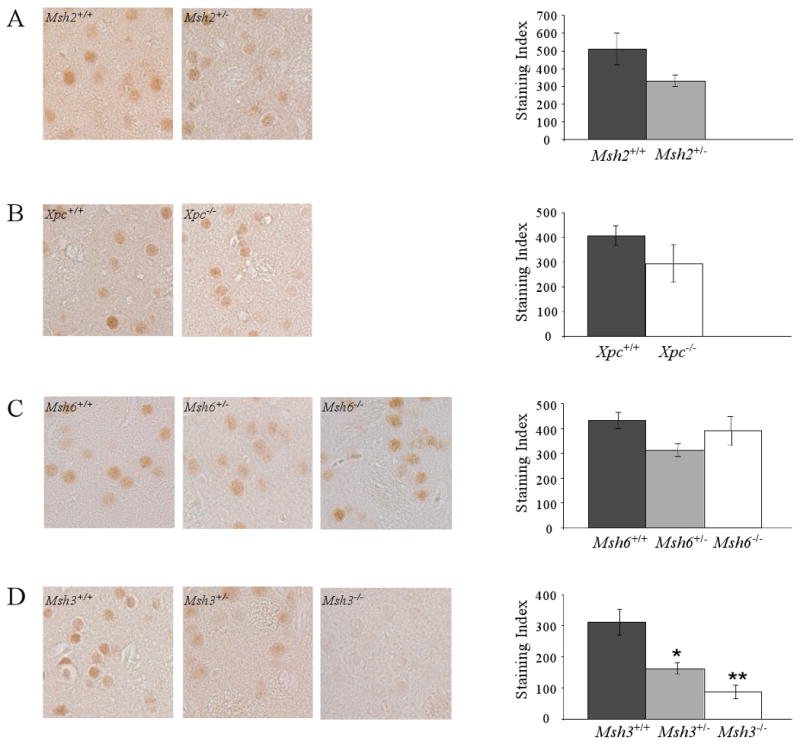
Modifying the length of the Huntington's disease (HD) CAG repeat, the major determinant of age of disease onset, is an attractive therapeutic approach. To explore this we are investigating mechanisms of intergenerational and somatic HD CAG repeat instability. Here, we have crossed HD CAG knock-in mice onto backgrounds deficient in mismatch repair genes, Msh3 and Msh6, to discern the effects on CAG repeat size and disease pathogenesis. We find that different mechanisms predominate in inherited and somatic instability, with Msh6 protecting against intergenerational contractions and Msh3 required both for increasing CAG length and for enhancing an early disease phenotype in striatum. Therefore, attempts to decrease inherited repeat size may entail a full understanding of Msh6 complexes, while attempts to block the age-dependent increases in CAG size in striatal neurons and to slow the disease process will require a full elucidation of Msh3 complexes and their function in CAG repeat instability.
Figures
References
-
- Andrew SE, Goldberg YP, Kremer B, Telenius H, Theilmann J, Adam S, Starr E, Squitieri F, Lin B, Kalchman MA, Graham RK, Hayden MR. The relationship between trinucleotide (CAG) repeat length and clinical features of Huntington's disease. Nat Genet. 1993;4:398–403. - PubMed
-
- Chang DK, Ricciardiello L, Goel A, Chang CL, Boland CR. Steady-state regulation of the human DNA mismatch repair system. J Biol Chem. 2000;275:18424–18431. - PubMed
-
- Cheo DL, Ruven HJ, Meira LB, Hammer RE, Burns DK, Tappe NJ, van Zeeland AA, Mullenders LH, Friedberg EC. Characterization of defective nucleotide excision repair in XPC mutant mice. Mut Res. 1997;374:1–9. - PubMed
-
- Duyao M, Ambrose C, Myers R, Novelletto A, Persichetti F, Frontali M, Folstein S, Ross C, Franz M, Abbott M, et al. Trinucleotide repeat length instability and age of onset in Huntington's disease. Nat Genet. 1993;4:387–392. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
