

FANCI phosphorylation functions as a molecular switch to turn on the Fanconi anemia pathway
- PMID: 18931676
- PMCID: PMC3293454
- DOI: 10.1038/nsmb.1504
FANCI phosphorylation functions as a molecular switch to turn on the Fanconi anemia pathway
Abstract
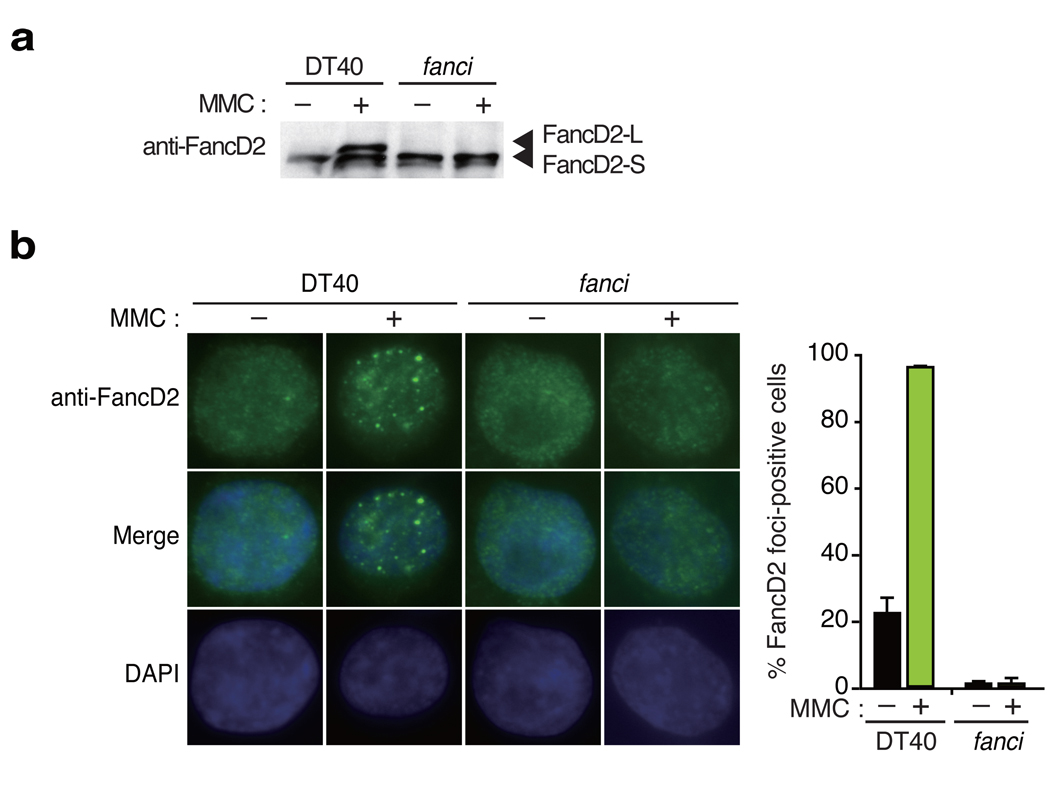
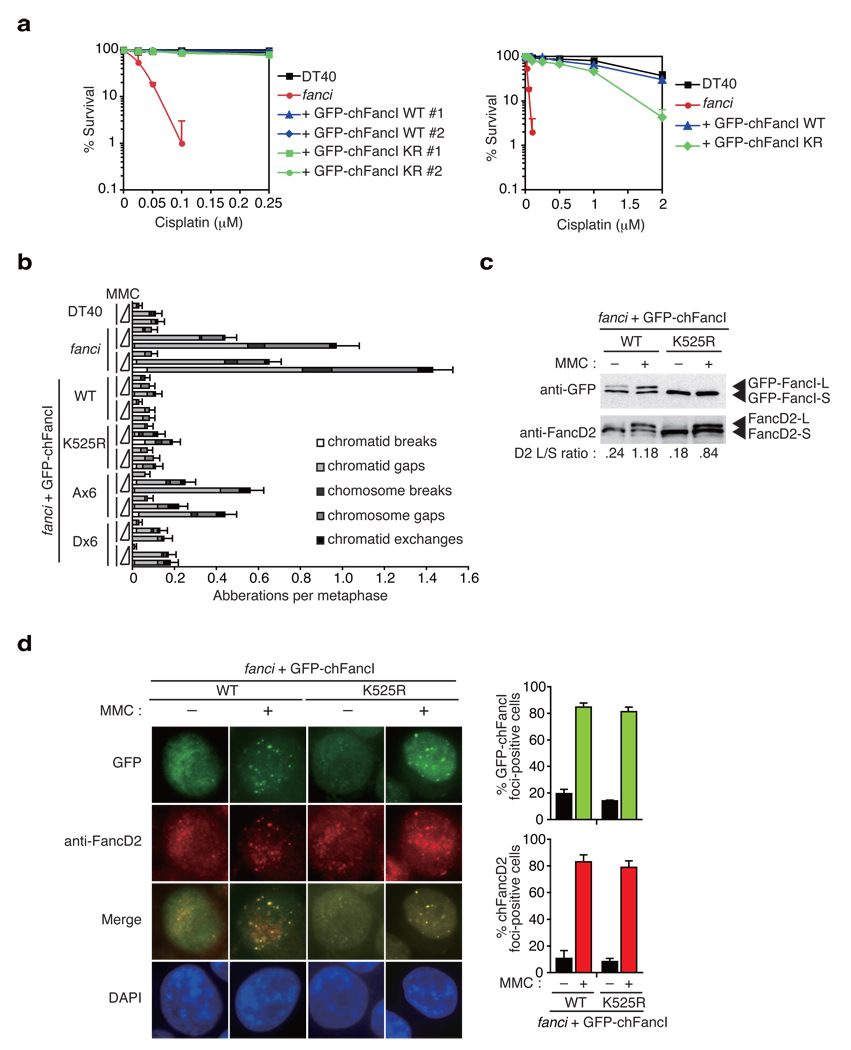
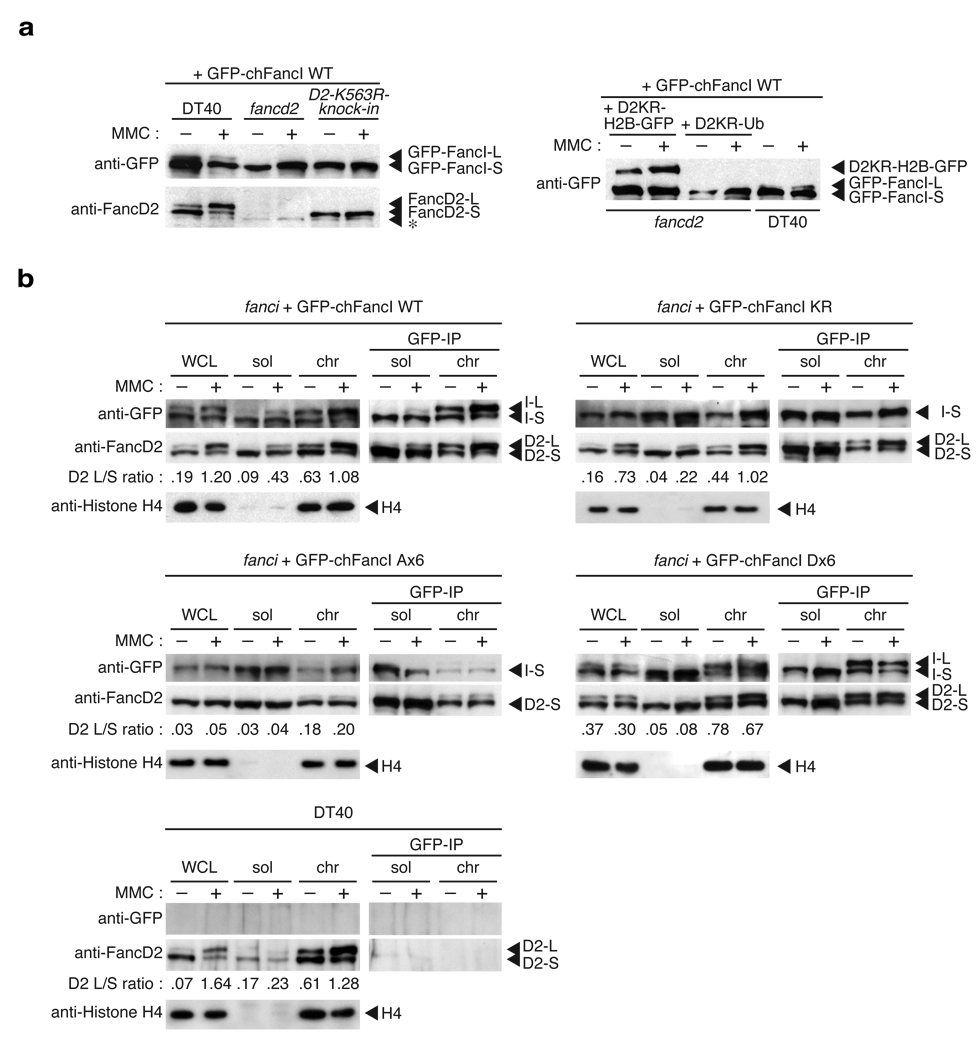
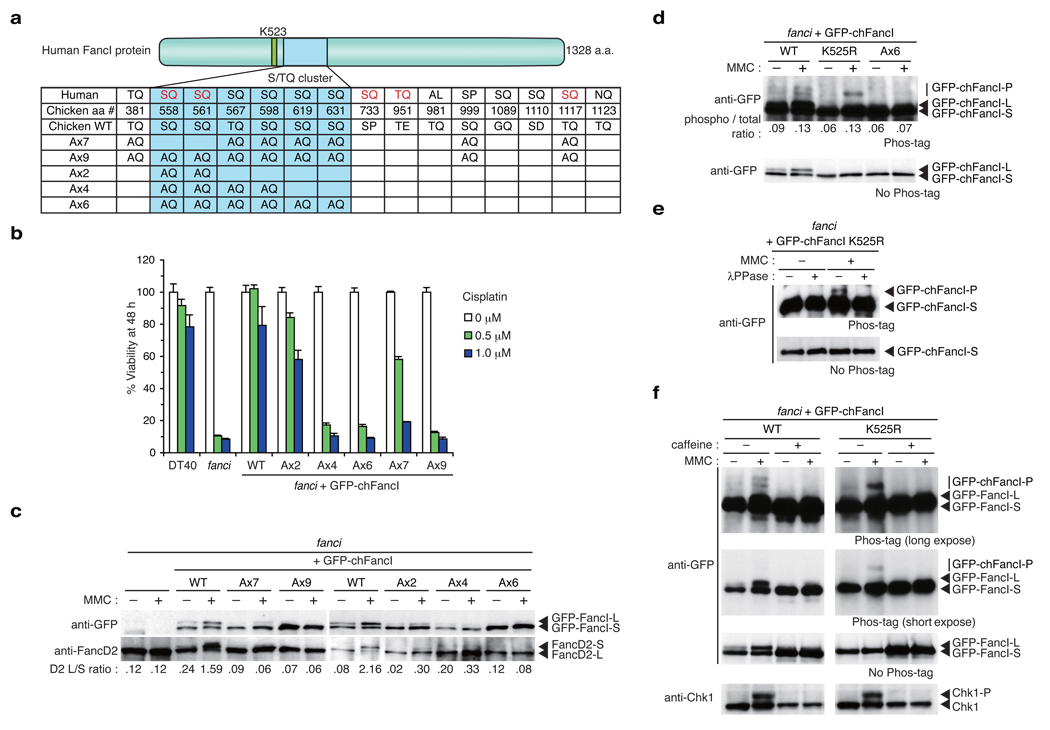
In response to DNA damage or replication fork stress, the Fanconi anemia pathway is activated, leading to monoubiquitination of FANCD2 and FANCI and their colocalization in foci. Here we show that, in the chicken DT40 cell system, multiple alanine-substitution mutations in six conserved and clustered Ser/Thr-Gln motifs of FANCI largely abrogate monoubiquitination and focus formation of both FANCI and FANCD2, resulting in loss of DNA repair function. Conversely, FANCI carrying phosphomimic mutations on the same six residues induces constitutive monoubiquitination and focus formation of FANCI and FANCD2, and protects against cell killing and chromosome breakage by DNA interstrand cross-linking agents. We propose that the multiple phosphorylation of FANCI serves as a molecular switch in activation of the Fanconi anemia pathway. Mutational analysis of putative phosphorylation sites in human FANCI indicates that this switch is evolutionarily conserved.
Figures
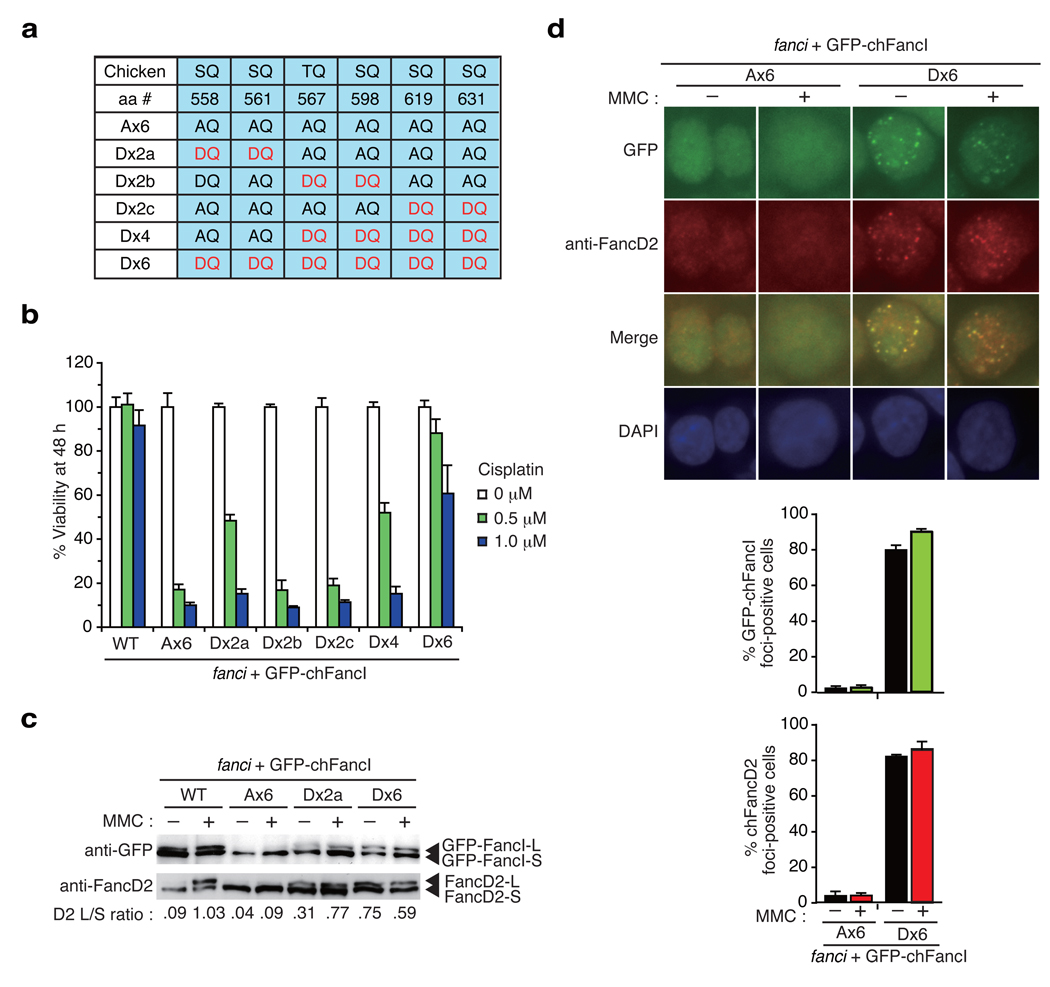
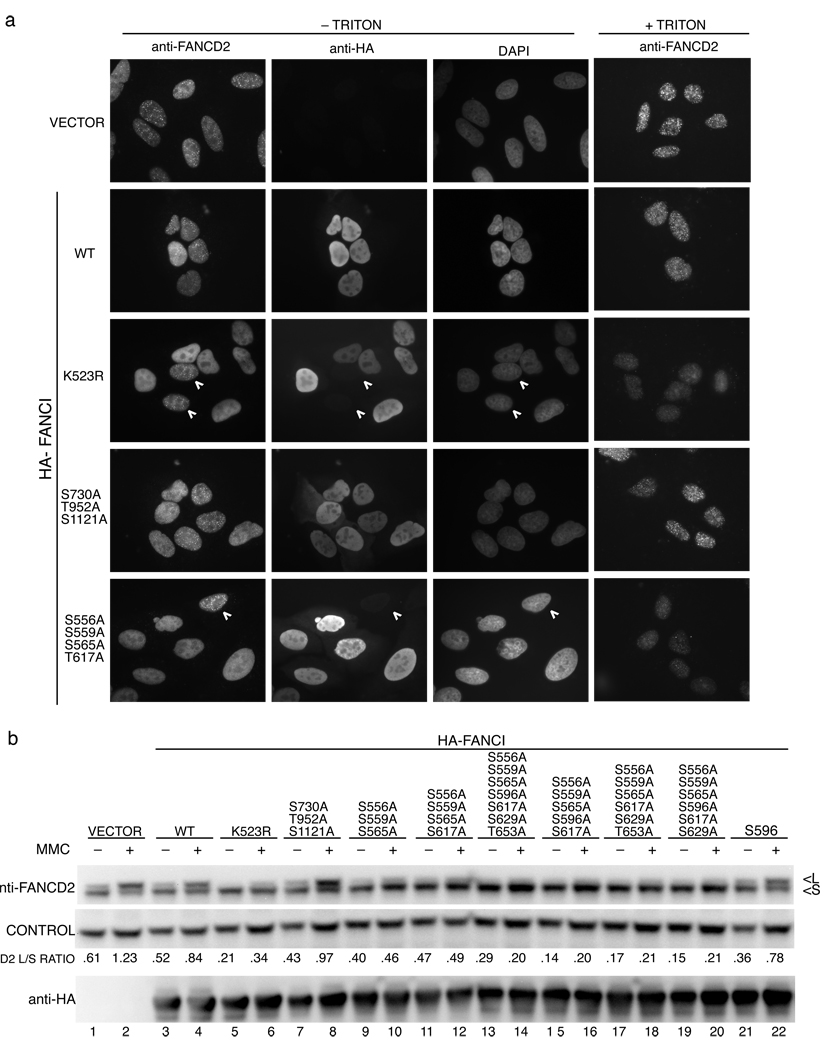
Comment in
-
A major switch for the Fanconi anemia DNA damage-response pathway.Nat Struct Mol Biol. 2008 Nov;15(11):1128-30. doi: 10.1038/nsmb1108-1128. Nat Struct Mol Biol. 2008. PMID: 18985065 No abstract available.
References
-
- Hoeijmakers JH. Genome maintenance mechanisms for preventing cancer. Nature. 2001;411:366–374. - PubMed
-
- Nijnik A, et al. DNA repair is limiting for haematopoietic stem cells during ageing. Nature. 2007;447:686–690. - PubMed
-
- Ito K, et al. Regulation of oxidative stress by ATM is required for self-renewal of haematopoietic stem cells. Nature. 2004;431:997–1002. - PubMed
-
- Rossi DJ, et al. Deficiencies in DNA damage repair limit the function of haematopoietic stem cells with age. Nature. 2007;447:725–729. - PubMed
-
- D'Andrea AD, Grompe M. The Fanconi anaemia/BRCA pathway. Nat. Rev. Cancer. 2003;3:23–34. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
