

Proteolytic cleavage in an endolysosomal compartment is required for activation of Toll-like receptor 9
- PMID: 18931679
- PMCID: PMC2735466
- DOI: 10.1038/ni.1669
Proteolytic cleavage in an endolysosomal compartment is required for activation of Toll-like receptor 9
Abstract
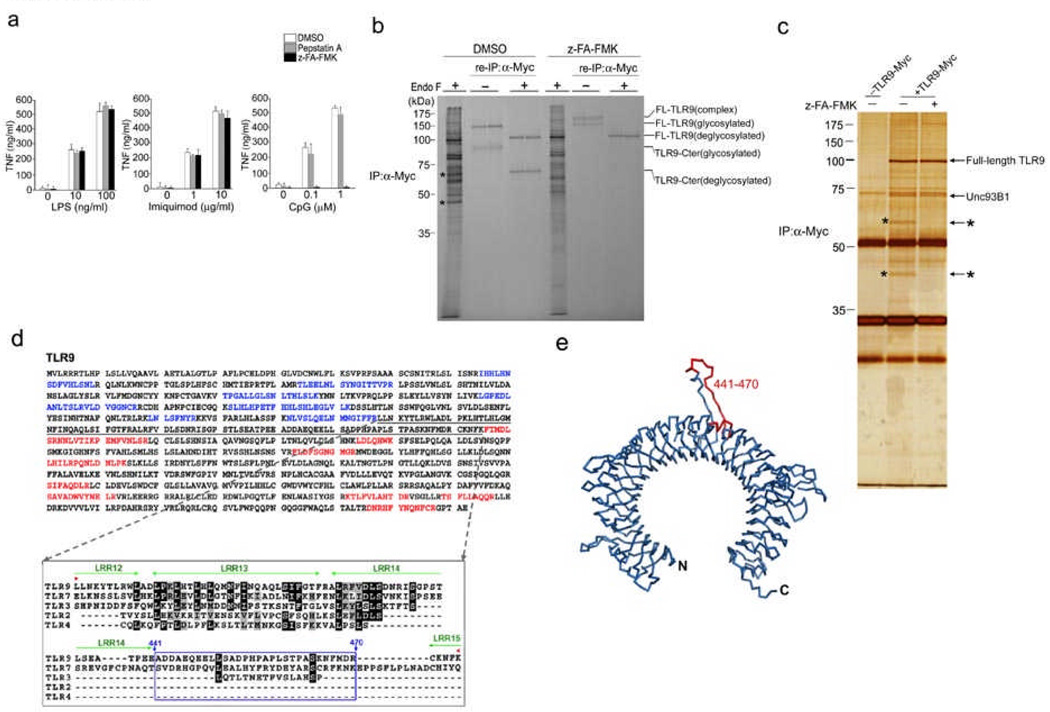
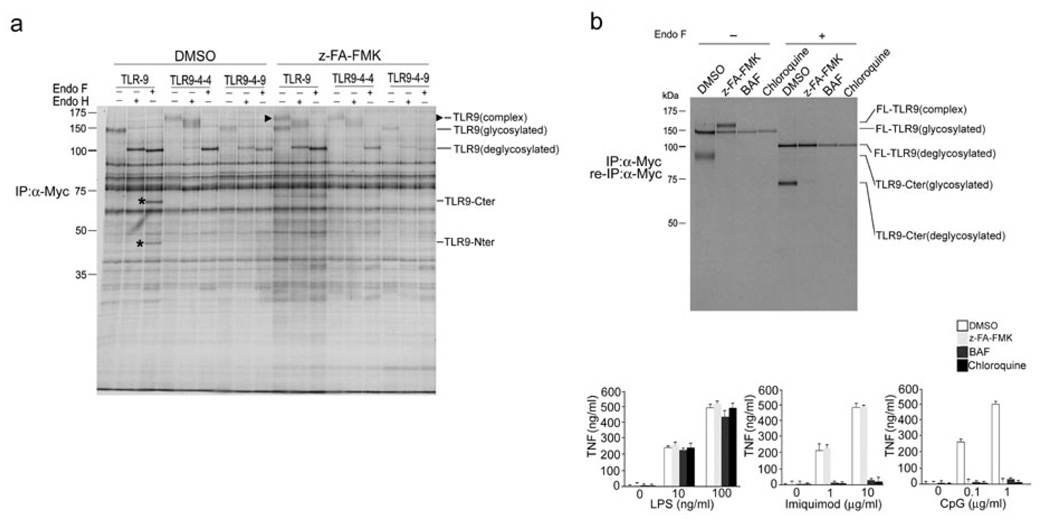
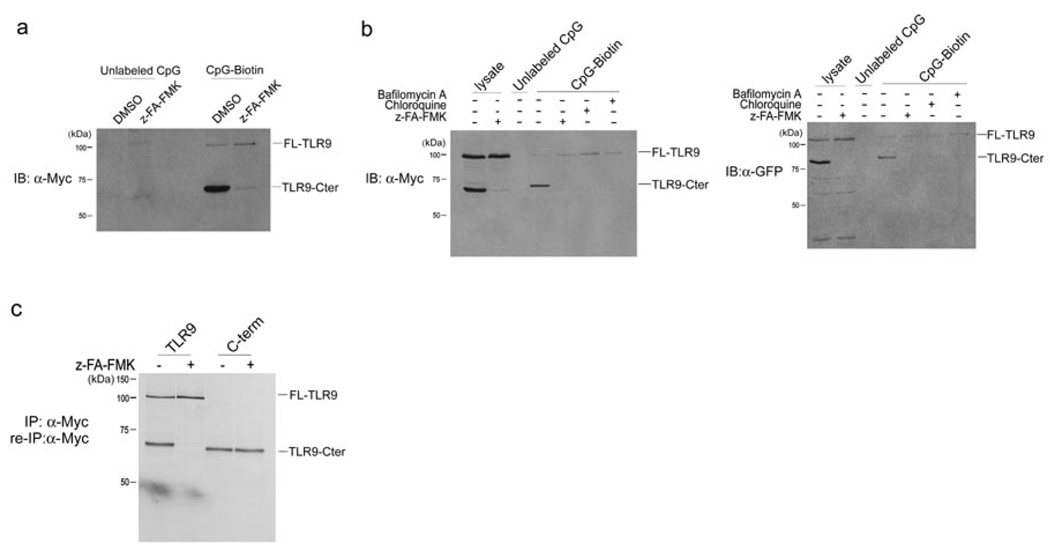
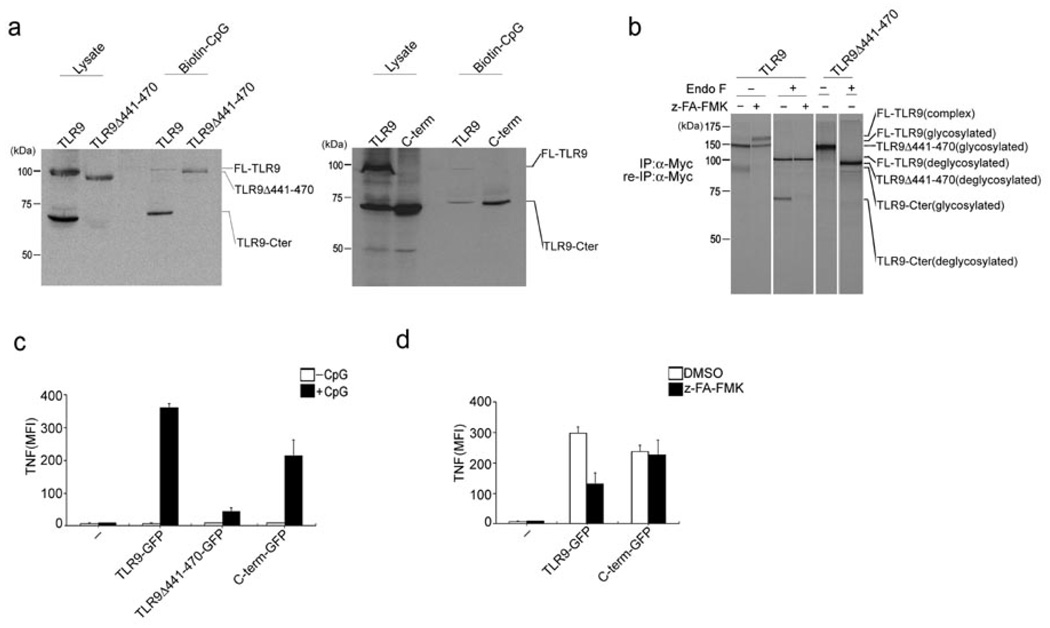
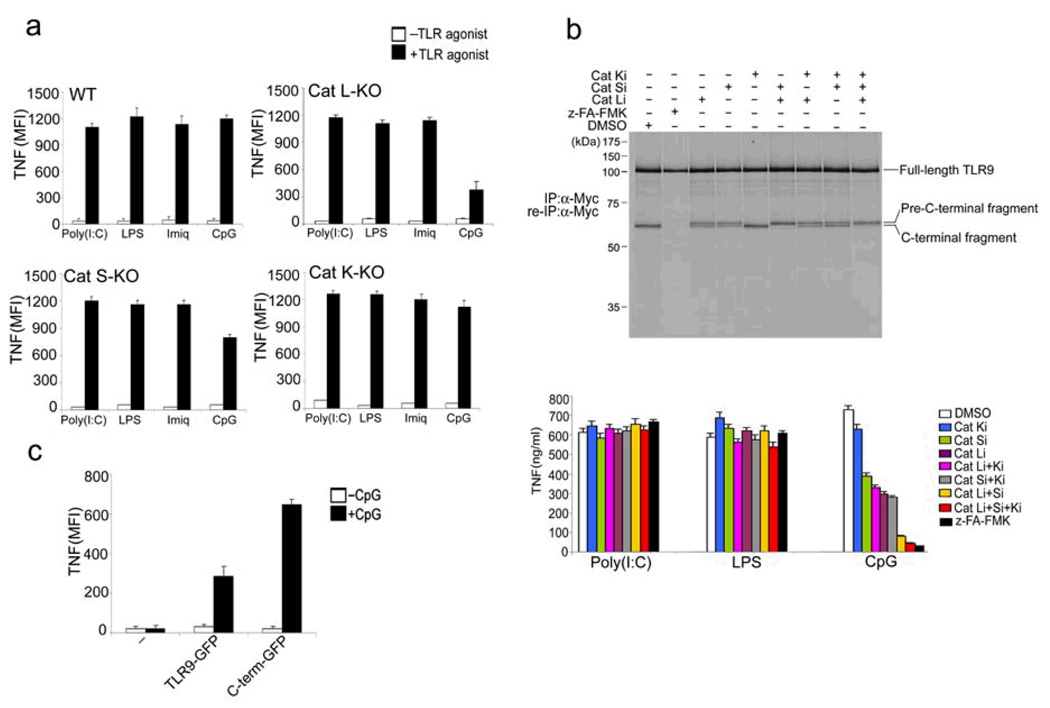
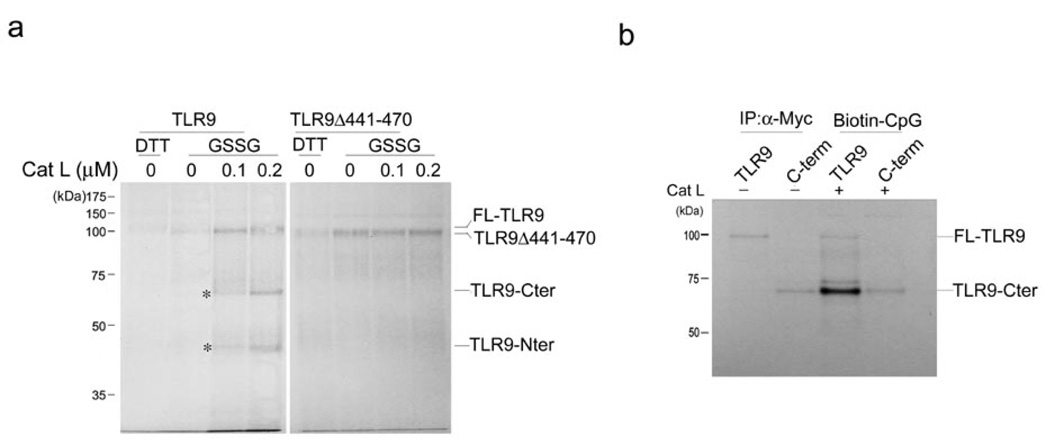
Toll-like receptors (TLRs) activate the innate immune system in response to pathogens. Here we show that TLR9 proteolytic cleavage is a prerequisite for TLR9 signaling. Inhibition of lysosomal proteolysis rendered TLR9 inactive. The carboxy-terminal fragment of TLR9 thus generated included a portion of the TLR9 ectodomain, as well as the transmembrane and cytoplasmic domains. This cleavage fragment bound to the TLR9 ligand CpG DNA and, when expressed in Tlr9(-/-) dendritic cells, restored CpG DNA-induced cytokine production. Although cathepsin L generated the requisite TLR9 cleavage products in a cell-free in vitro system, several proteases influenced TLR9 cleavage in intact cells. Lysosomal proteolysis thus contributes to innate immunity by facilitating specific cleavage of TLR9.
Figures
References
-
- Takeda K, Kaisho T, Akira S. Toll-like receptors. Annu Rev Immunol. 2003;21:335–376. - PubMed
-
- Janeway CA, Jr, Medzhitov R. Innate immune recognition. Annu Rev Immunol. 2002;20:197–216. - PubMed
-
- Kawai T, Akira S. TLR signaling. Cell Death Differ. 2006;13:816–825. - PubMed
-
- Takeda K, Akira S. Toll-like receptors in innate immunity. Int Immunol. 2005;17:1–14. - PubMed
-
- Poltorak A, et al. Defective LPS signaling in C3H/HeJ and C57BL/10ScCr mice: mutations in Tlr4 gene. Science. 1998;282:2085–2028. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
