

doi: 10.1038/ng.235.
Epub 2008 Oct 19.
Structure and function of a transcriptional network activated by the MAPK Hog1
Affiliations
- PMID: 18931682
- PMCID: PMC2825711
- DOI: 10.1038/ng.235
Item in Clipboard
Structure and function of a transcriptional network activated by the MAPK Hog1
Nat Genet.
2008 Nov.
Abstract
Cells regulate gene expression using a complex network of signaling pathways, transcription factors and promoters. To gain insight into the structure and function of these networks, we analyzed gene expression in single- and multiple-mutant strains to build a quantitative model of the Hog1 MAPK-dependent osmotic stress response in budding yeast. Our model reveals that the Hog1 and general stress (Msn2/4) pathways interact, at both the signaling and promoter level, to integrate information and create a context-dependent response. This study lays out a path to identifying and characterizing the role of signal integration and processing in other gene regulatory networks.
Figures
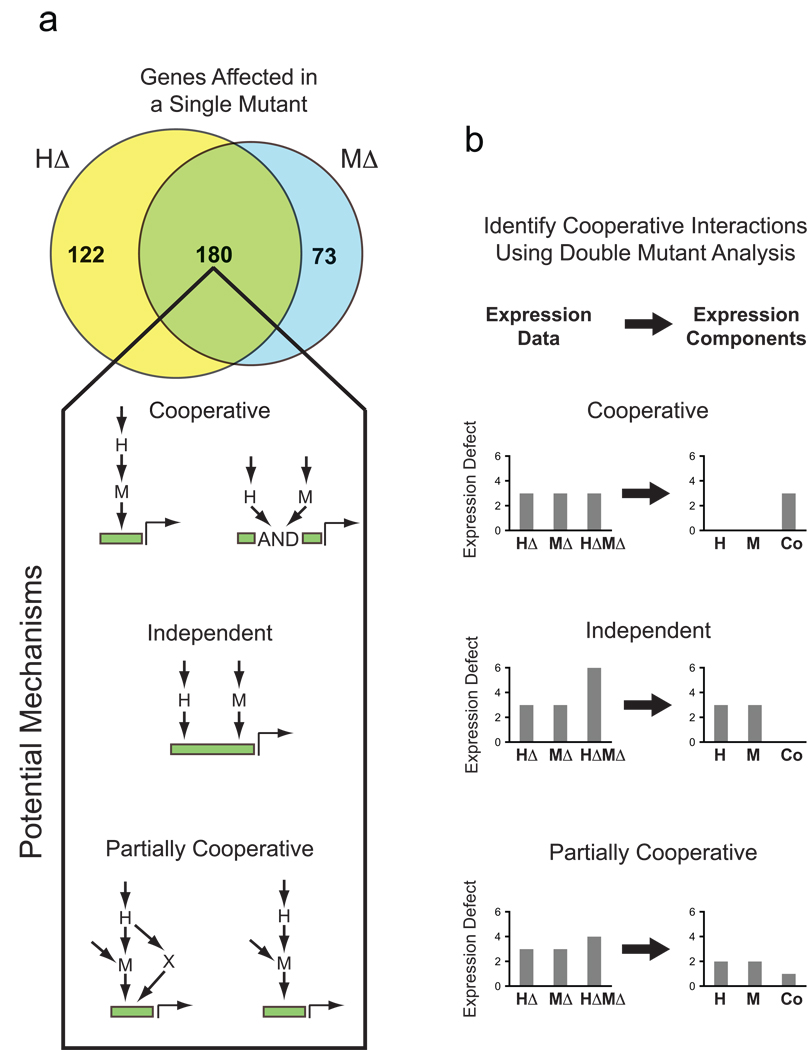
Single and double mutant analysis of gene expression. (A) Venn diagram summarizing the overlap in the number of genes with a >2-fold (log2=1) defect in gene expression in the hog1Δ (HΔ) and msn2Δ msn4Δ (MΔ) mutants, following salt induction. Wiring diagrams indicate the possible ways factors H and M can interact to regulate expression of overlapping sets of genes. (B) Schematic illustrating the application of the double mutant approach to analyzing transcriptional network structure and function (see text for details).
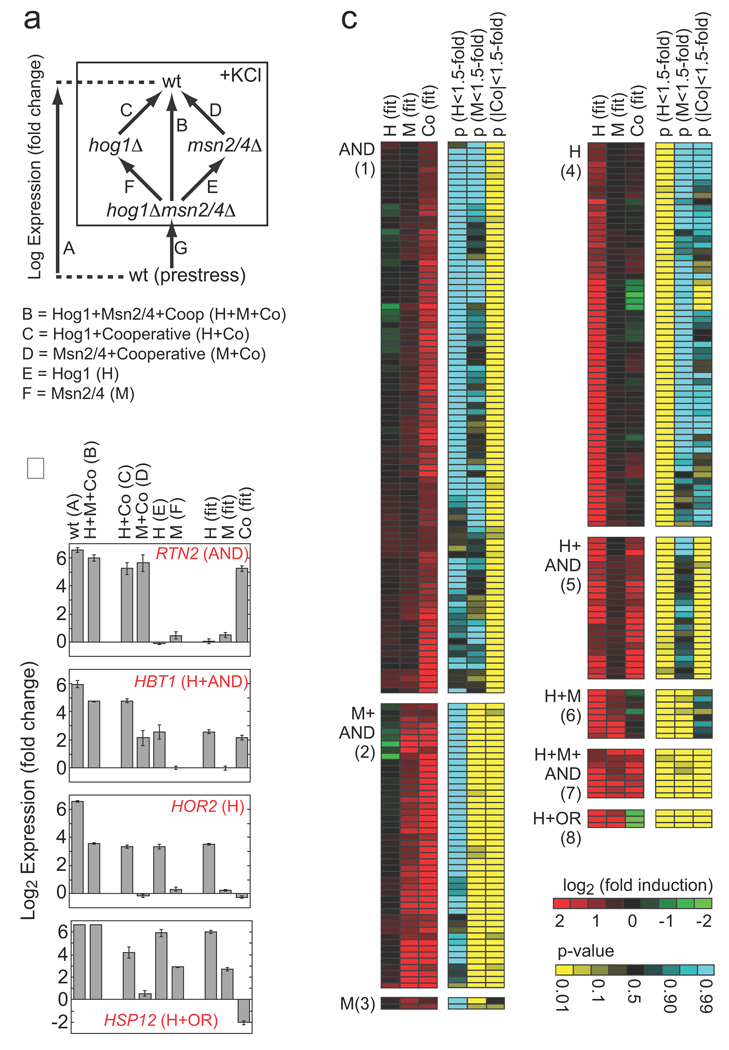
Role of Hog1 and Msn2/4 in osmotic stress-dependent gene induction. (A) Schema describing the experiments and equations used to break the influence of Hog1 and Msn2/4 into components. Each arrow represents a single microarray (measured in triplicate) comparing gene expression in two strains. The equations listed below the diagram describe the relationship between the data from each measurement and the underlying expression components. Note here that expression is in Log terms and thus an OR gate is manifest as a negative cooperative component equal to the H or M component (see Supplement). (B) Sample data for four genes showing the errors associated with the microarray measurements and expression component values. (C) Heat map showing the best-fit value of the expression components (red/green), and their statistical significance (yellow/blue), for all genes that are upregulated more than 3-fold in response to hyperosmotic stress, by Hog1 or Msn2/4 (>2-fold). Each row of six columns shows the data for a single gene. Genes are placed into groups (1–8) and labeled according to the combination of expression components (p<0.05 cut-off) that influence their induction (AND = +Co, OR = −Co). Data is not shown for 15 genes that are induced in the wild-type strain (>3 fold) by Hog1 and/or Msn2/4 (>2-fold) but have no significant expression component. See Tables S2 and S3 for full data.
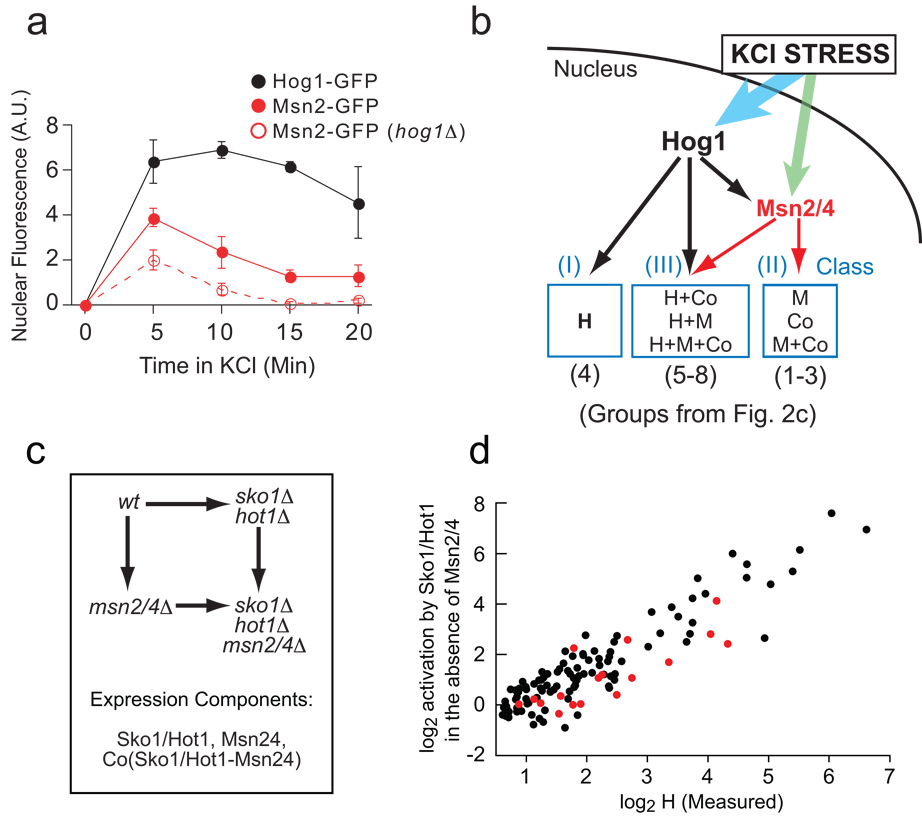
Mechanism of Hog1-dependent gene activation. (A) Hog1 promotes the nuclear import of Msn2/4 in hyper-osmotic stress. Fluorescence microscopy was used to measure the relative nuclear concentration of Hog1-GFP or Msn2-GFP, in live cells, after exposure to 0.4 M KCl. Each time-point shows the average and standard deviation from three replicate experiments, each involving 100 or more cells (Table S4). (B) Model of the Hog1 transcriptional network, explaining the expression component data found in Fig. 2 (see text for details). (C) Schema describing the experiments and equations used to break the influence of Sko1, Hot1 and Msn2/4 into components (as in Fig. 2a). (D) Correlation between the level of induction measured for Hog1 alone (H component, Fig. 2) and that from Sko1/ Hot1 in the absence of Msn2/4 (Sko1/Hot1 component, cycle part Fig. 3c) plus Sko1 repression in YEPD data (Table S1 and Supplement).
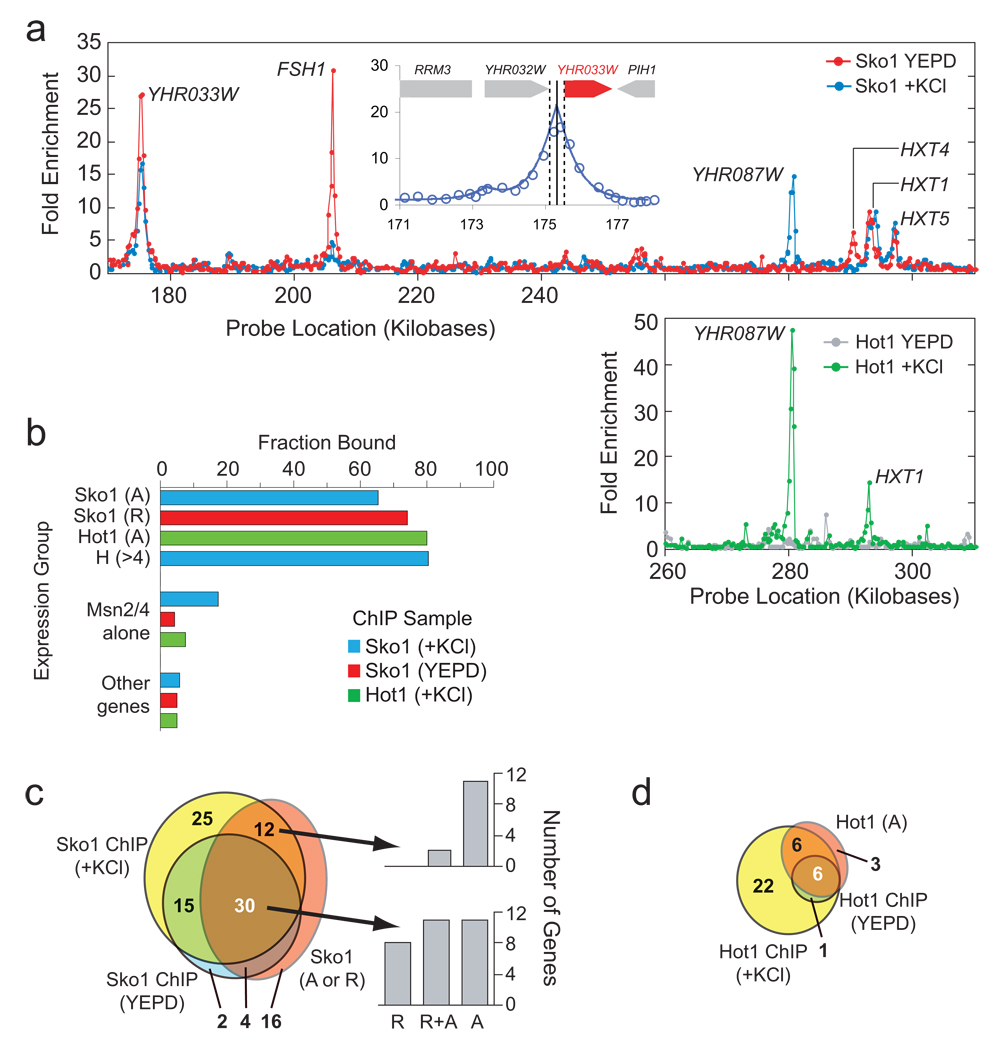
ChIP analysis of Sko1 and Hot1 binding sites (A) Sample raw data for Sko1 (upper panel) and Hot1 (lower panel) for a region of chromosome 8 (approximately 1% of the genome). Each data point shows the Cy5/Cy3 ratio for one probe on the array. The inset shows an example of a fit of the data to the peak shape model used to analyze the data (see Methods and Supplement). The solid line shows the best-fit prediction of the binding site position, while the dotted lines show the 99% confidence interval. The ChIP data is listed in Tables S2 and S3. (B) Overlap of ChIP and expression data. The target genes shown in Fig. S5c for Sko1, Hot1 and Msn2/4 alone (p<0.05) were compared to the target genes identified in the ChIP analysis from the peak fitting (p<0.05). In the case of Sko1 (+KCl) the p-value was relaxed to 0.058 since we found significantly better coverage at this value. This is likely due to a lower binding affinity of Sko1 to genes that are only bound in stress conditions (and thus a lower peak height/significance). (C) Venn diagram showing the overlap between ChIP data (p<0.05) and expression data (p<0.058) for Sko1. The number of binding sites at genes without significant Sko1 induction and/or repression, is adjusted for the expected number of false positives. The bar graphs show the number of genes that are repressed (R), repressed and activated (R+A) or just activated, for genes where there is both significant binding and expression data. (D) Venn diagram showing the overlap between ChIP data (p<0.05) and expression data (p<0.05) for Hot1. Again here the number of binding sites at genes without significant Hot1 induction is corrected for the number of false positives expected.
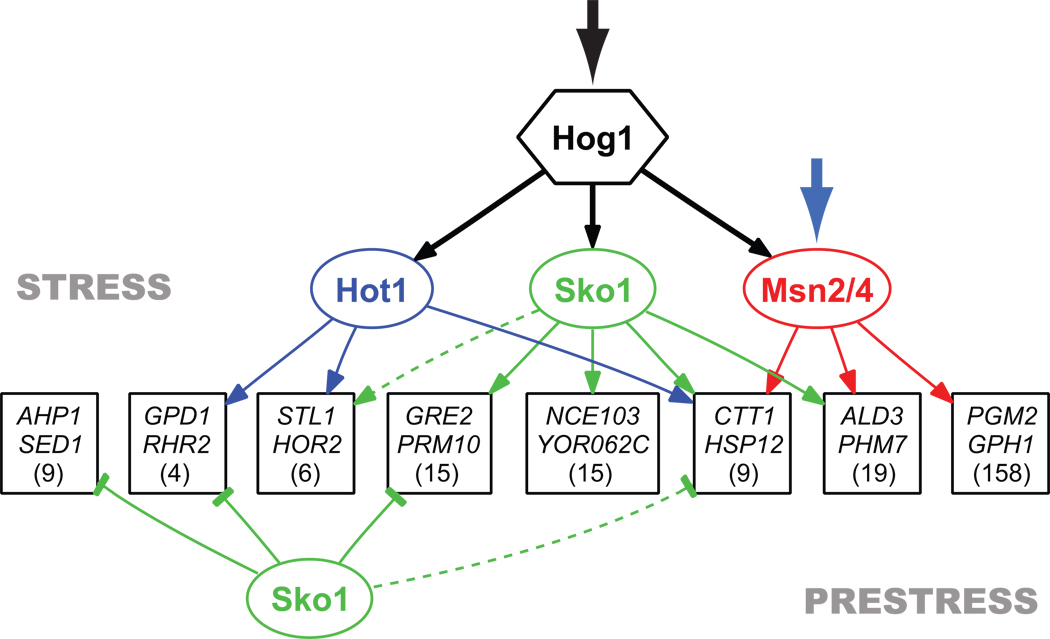
Structure of the transcriptional network activated by the MAPK Hog1. Genes are grouped based on common regulatory mechanisms (denoted by a box with the names of two sample genes) and only shown if two or more genes have the same connections as determined by expression and confirmed by ChIP. Broken lines indicate interactions that that exist for only part of a group. The number in each box refers to the number of genes in a group based on expression data alone. To simplify the figure silent binding events are not shown and there is no representation of cooperativity at the promoter level. All of the values describing the network are listed in Table S2.
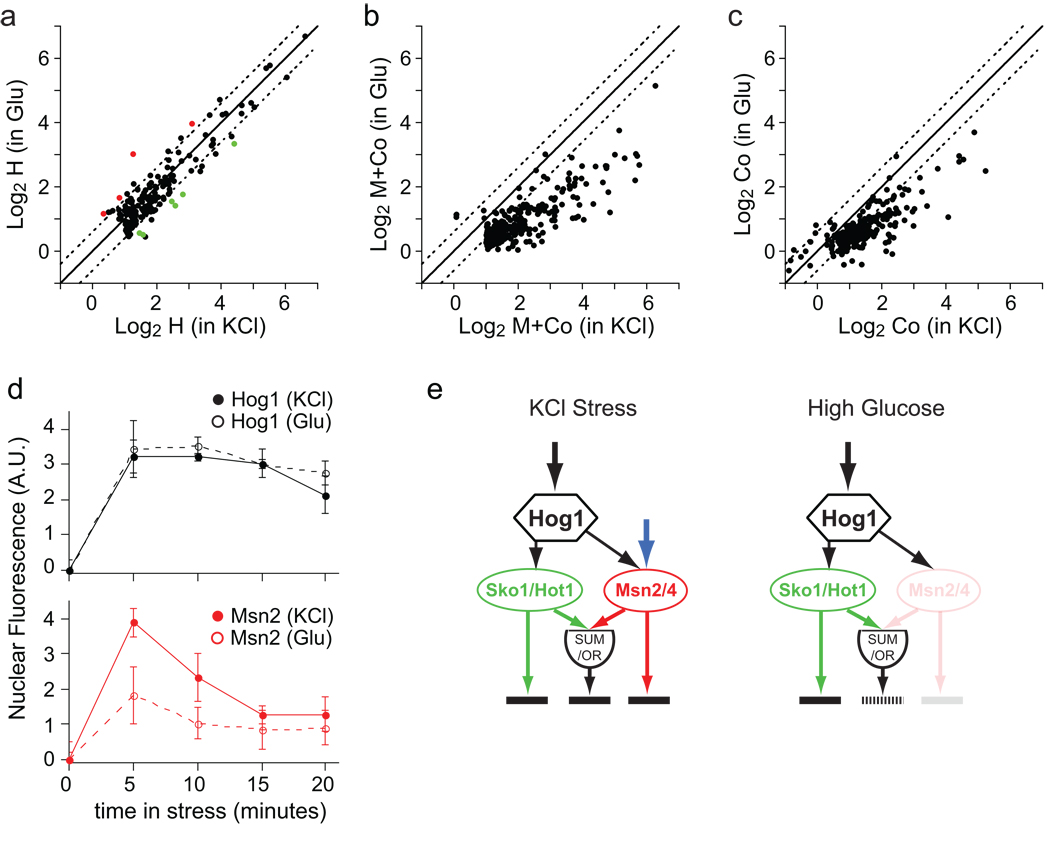
Context-dependent gene activation by the Hog1-Msn2/4 network. (A) Plot comparing the H component in KCl stress (0.4 M) and glucose stress (0.8 M). Each point shows the data for a single gene; colored red if (HGlu-HKCl)<1.5, p<0.05; green if (HKCl-HGlu)<1.5, p<0.05); and black if there is no significant change. The solid and broken lines show the values expected for perfect correlation and a ±1.5-fold difference, respectively. Data is shown for all genes with a significant H component (H<1.5-fold, p<0.01) in KCl or glucose (n=170). (B) Plot comparing the total influence of Msn2/4 (M component + Co component), in osmotic stress due to 0.4 M KCl (X-axis) or 0.8 M glucose (Y-axis). Gene selection (M+Co<1.5-fold, p<0.01; n=280) and lines are as in (A). (C) Plot comparing the cooperative influence of Hog1 and Msn2/4 (Co component) on gene expression, in osmotic stress due to 0.4 M KCl or 0.8 M glucose. The lines are as in (C) and the genes are those shown in (B). (D) Time-course of Hog1 and Msn2 nuclear import during KCl and glucose stress (as described for Fig. 3a). (E) Model of the Hog1-Msn2/4 network in KCl (left panel) and high glucose (right panel).
References
-
- Alon U. An introduction to systems biology : design principles of biological circuits. Boca Raton, FL: Chapman & Hall/CRC; 2007.
-
- Davidson EH. The regulatory genome : gene regulatory networks in development and evolution. Burlington, MA: Academic; 2006.
-
- Hu Z, Killion PJ, Iyer VR. Genetic reconstruction of a functional transcriptional regulatory network. Nat Genet. 2007;39:683–687. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
