

Novel proteolytic processing of the ectodomain of the zinc transporter ZIP4 (SLC39A4) during zinc deficiency is inhibited by acrodermatitis enteropathica mutations
- PMID: 18936158
- PMCID: PMC2612479
- DOI: 10.1128/MCB.00963-08
Novel proteolytic processing of the ectodomain of the zinc transporter ZIP4 (SLC39A4) during zinc deficiency is inhibited by acrodermatitis enteropathica mutations
Abstract
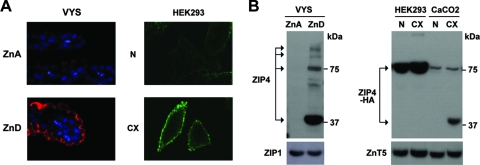
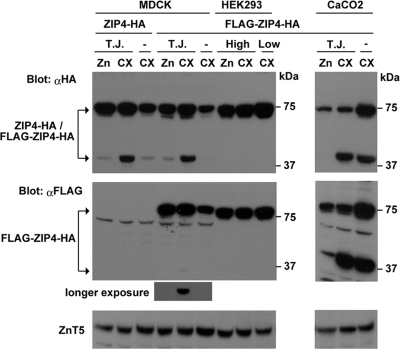
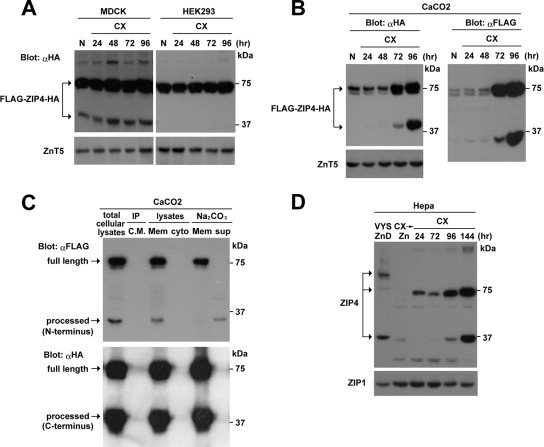
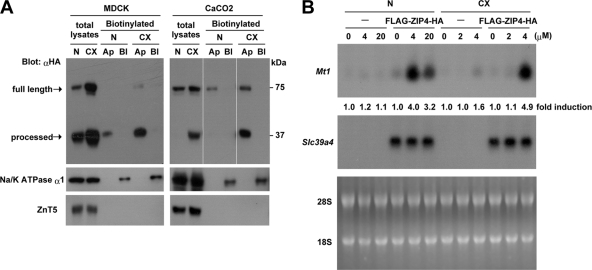
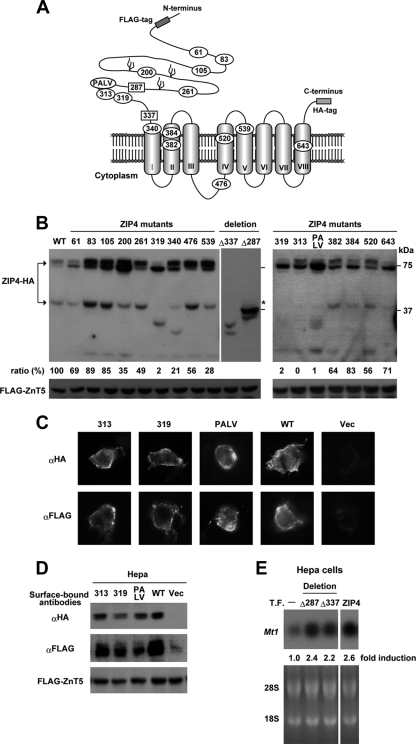
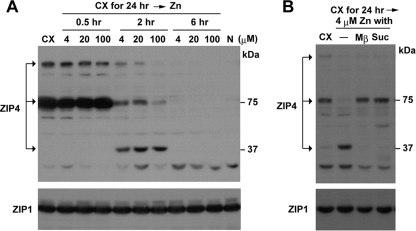
The zinc transporter ZIP4 (SLC39A4) is mutated in humans with the rare, autosomal recessive genetic disease acrodermatitis enteropathica. In mice, this gene is essential during early embryonic development. ZIP4 is dynamically regulated by multiple posttranscriptional mechanisms, and studies of mouse ZIP4 reported herein reveal that the ectodomain, the extracellular amino-terminal half of the protein, is proteolytically removed during prolonged zinc deficiency while the remaining eight-transmembrane carboxyl-terminal half of the protein is accumulated on the plasma membrane as an abundant form of ZIP4. This novel ZIP4 processing occurs in vivo in the intestine and visceral endoderm, in mouse Hepa cells that express the endogenous Slc39a4 gene and in transfected MDCK and CaCo2 cells, but not HEK293 cells. In transfected MDCK and CaCo2 cells, the ectodomain accumulated and remained associated with membranes when zinc was deficient. ZIP4 cleavage was attenuated by inhibitors of endocytosis, which suggests that the processed protein is recycled back to the plasma membrane and that the ectodomain may be internalized. Ectodomain cleavage is inhibited by acrodermatitis enteropathica mutations near a predicted metalloproteinase cleavage site which is also essential for proper ectodomain cleavage, and overexpression of processed ZIP4 or ZIP4 with ectodomain truncations rendered the mouse Mt1 gene hypersensitive to zinc. These finding suggest that the processing of ZIP4 may represent a significant regulatory mechanism controlling its function.
Figures

References
-
- Chen, E. I., S. J. Kridel, E. W. Howard, W. Li, A. Godzik, and J. W. Smith. 2002. A unique substrate recognition profile for matrix metalloproteinase-2. J. Biol. Chem. 2774485-4491. - PubMed
-
- Dufner-Beattie, J., Y. M. Kuo, J. Gitschier, and G. K. Andrews. 2004. The adaptive response to dietary zinc in mice involves the differential cellular localization and zinc-regulation of the zinc transporters ZIP4 and ZIP5. J. Biol. Chem. 27949082-49090. - PubMed
-
- Dufner-Beattie, J., S. J. Langmade, F. Wang, D. Eide, and G. K. Andrews. 2003. Structure, function, and regulation of a subfamily of mouse zinc transporter genes. J. Biol. Chem. 27850142-50150. - PubMed
-
- Dufner-Beattie, J., F. Wang, Y. M. Kuo, J. Gitschier, D. Eide, and G. K. Andrews. 2003. The acrodermatitis enteropathica gene ZIP4 encodes a tissue-specific, zinc-regulated zinc transporter in mice. J. Biol. Chem. 27833474-33481. - PubMed
-
- Dufner-Beattie, J., B. P. Weaver, J. Geiser, M. Bilgen, M. Larson, W. Xu, and G. K. Andrews. 2007. The mouse acrodermatitis gene Slc39a4 (ZIP4) is essential for development and heterozygosity causes hypersensitivity to zinc deficiency. Hum. Mol. Genet. 161391-1399. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases