

The PmrA/PmrB two-component system of Legionella pneumophila is a global regulator required for intracellular replication within macrophages and protozoa
- PMID: 18936184
- PMCID: PMC2612241
- DOI: 10.1128/IAI.01081-08
The PmrA/PmrB two-component system of Legionella pneumophila is a global regulator required for intracellular replication within macrophages and protozoa
Abstract
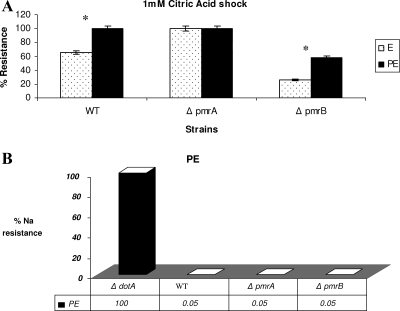
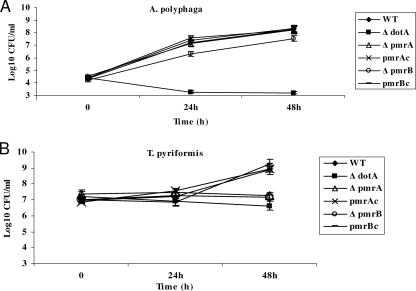
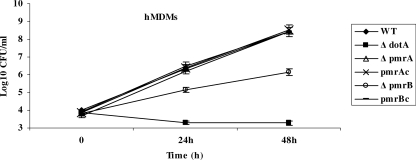
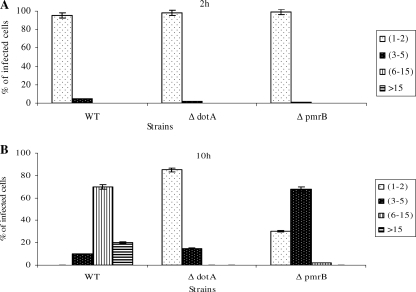
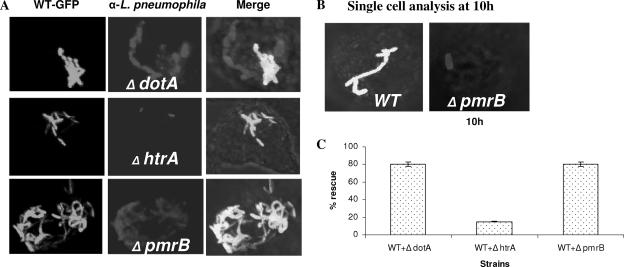
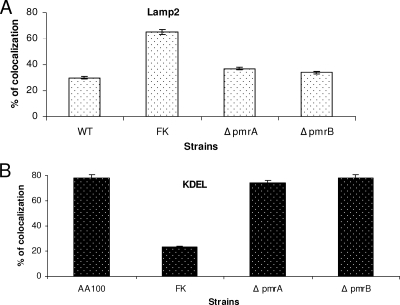
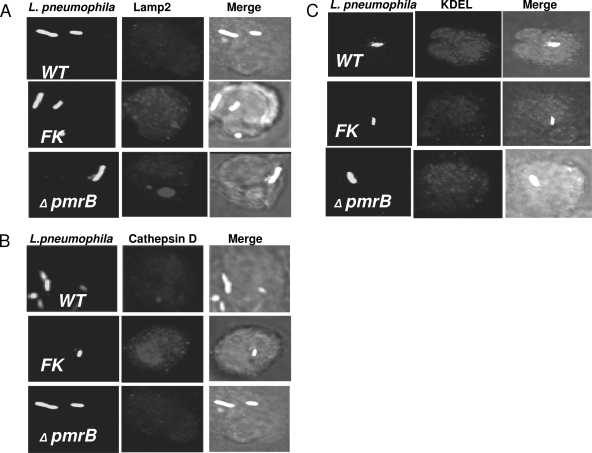
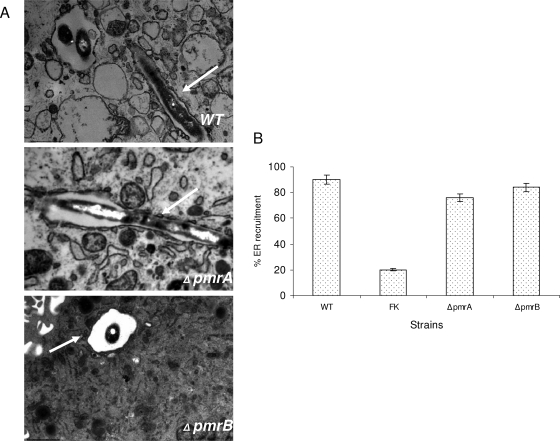
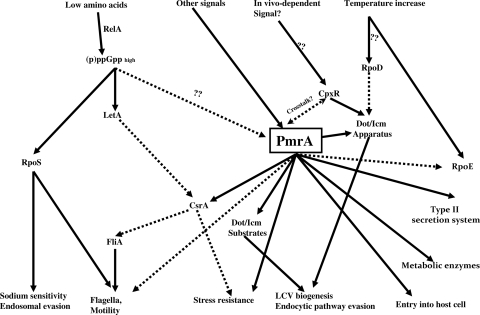
To examine the role of the PmrA/PmrB two-component system (TCS) of Legionella pneumophila in global gene regulation and in intracellular infection, we constructed pmrA and pmrB isogenic mutants by allelic exchange. Genome-wide microarray gene expression analyses of the pmrA and pmrB mutants at both the exponential and the postexponential phases have shown that the PmrA/PmrB TCS has a global effect on the expression of 279 genes classified into nine groups of genes encoding eukaryotic-like proteins, Dot/Icm apparatus and secreted effectors, type II-secreted proteins, regulators of the postexponential phase, stress response genes, flagellar biosynthesis genes, metabolic genes, and genes of unknown function. Forty-one genes were differentially regulated in the pmrA or pmrB mutant, suggesting a possible cross talk with other TCSs. The pmrB mutant is more sensitive to low pH than the pmrA mutant and the wild-type strain, suggesting that acidity may trigger this TCS. The pmrB mutant exhibits a significant defect in intracellular proliferation within human macrophages, Acanthamoeba polyphaga, and the ciliate Tetrahymena pyriformis. In contrast, the pmrA mutant is defective only in the ciliate. Despite the intracellular growth defect within human macrophages, phagosomes harboring the pmrB mutant exclude late endosomal and lysosomal markers and are remodeled by the rough endoplasmic reticulum. Similar to the dot/icm mutants, the intracellular growth defect of the pmrB mutant is totally rescued in cis within communal phagosomes harboring the wild-type strain. We conclude that the PmrA/PmrB TCS has a global effect on gene expression and is required for the intracellular proliferation of L. pneumophila within human macrophages and protozoa. Differences in gene regulation and intracellular growth phenotypes between the pmrA and pmrB mutant suggests a cross talk with other TCSs.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
