

Increased levels of superoxide and H2O2 mediate the differential susceptibility of cancer cells versus normal cells to glucose deprivation
- PMID: 18937644
- PMCID: PMC2678564
- DOI: 10.1042/BJ20081258
Increased levels of superoxide and H2O2 mediate the differential susceptibility of cancer cells versus normal cells to glucose deprivation
Abstract
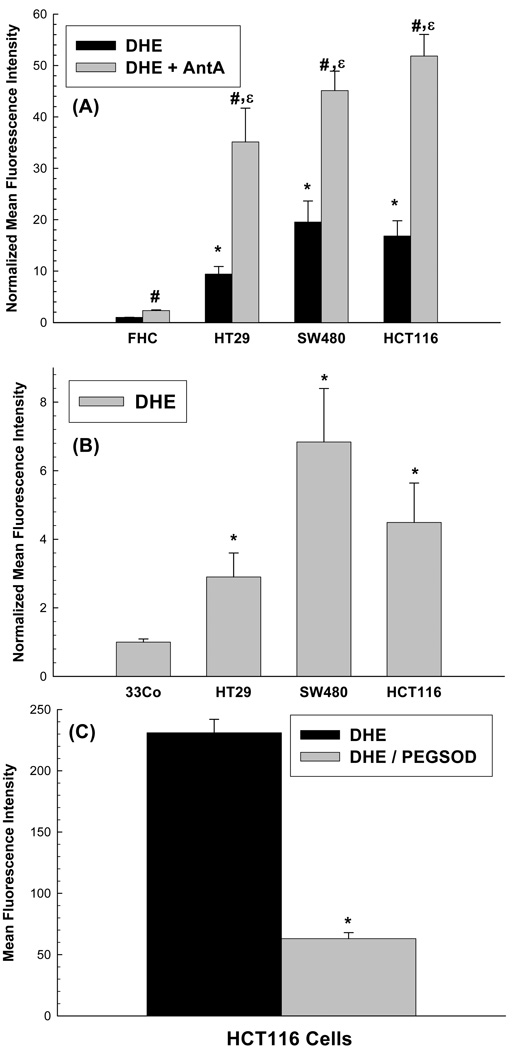
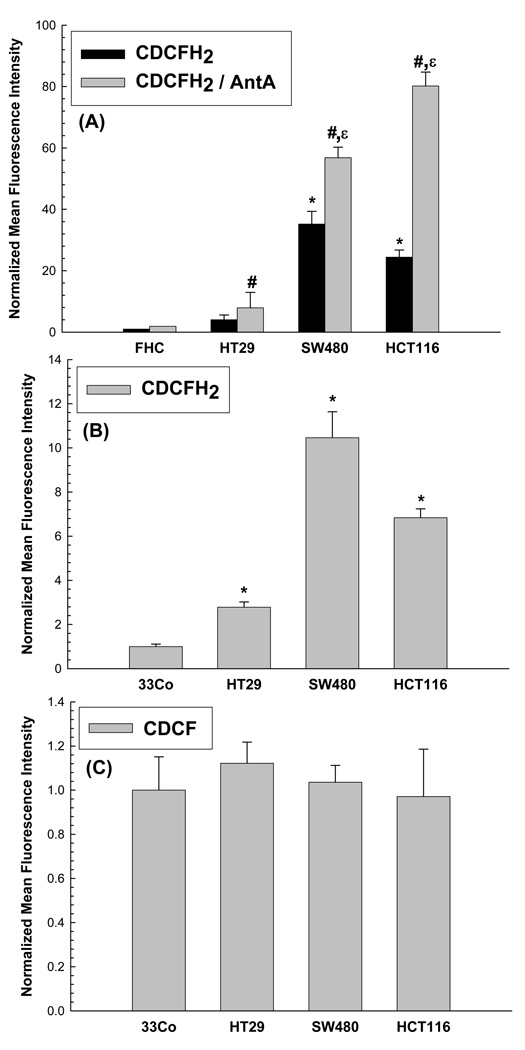
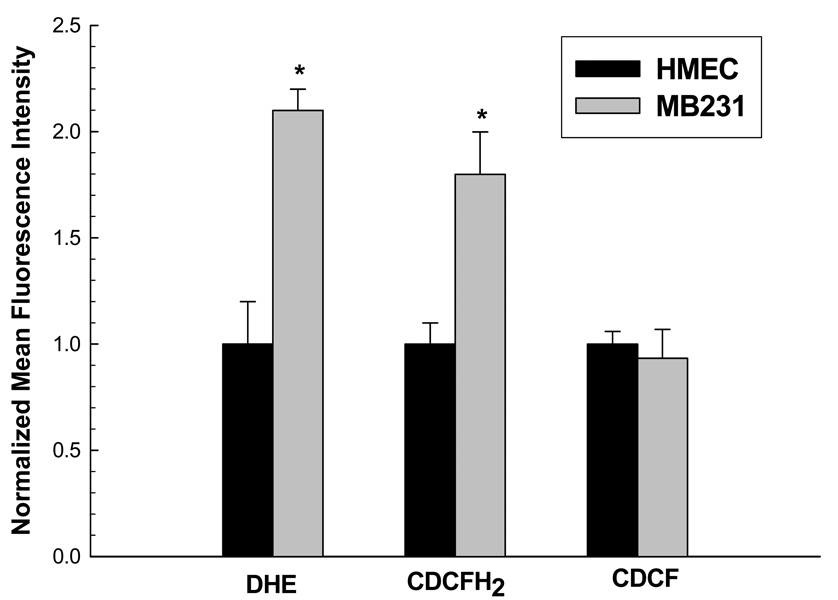
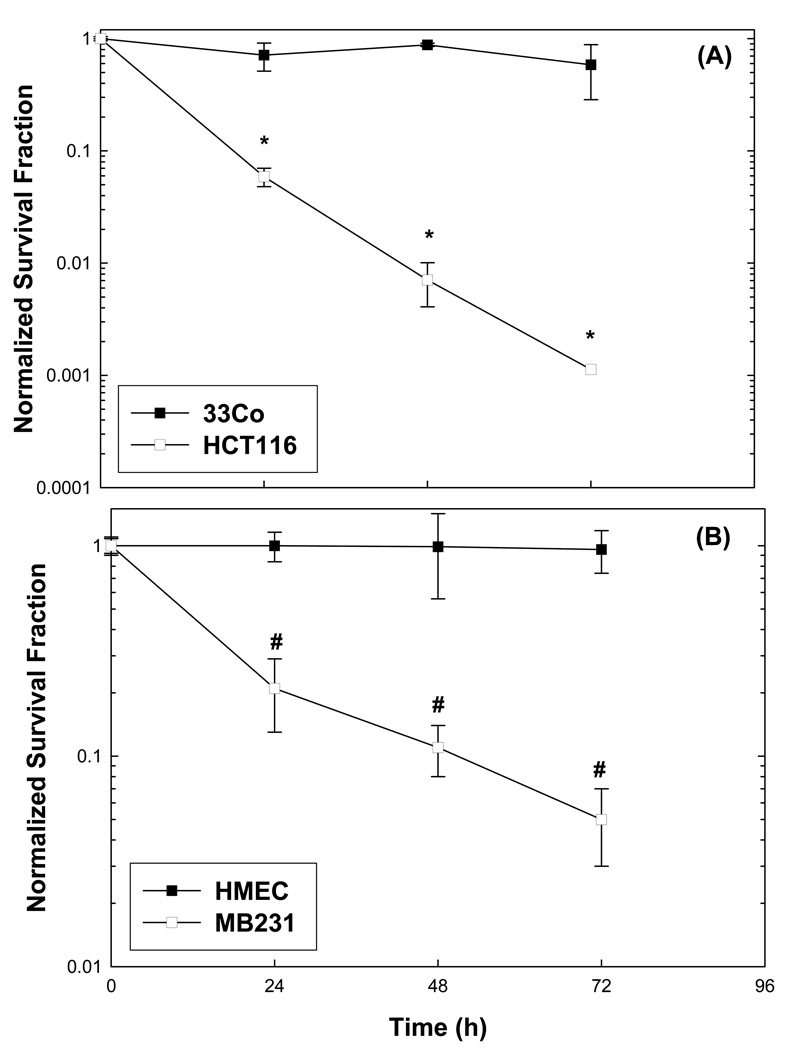
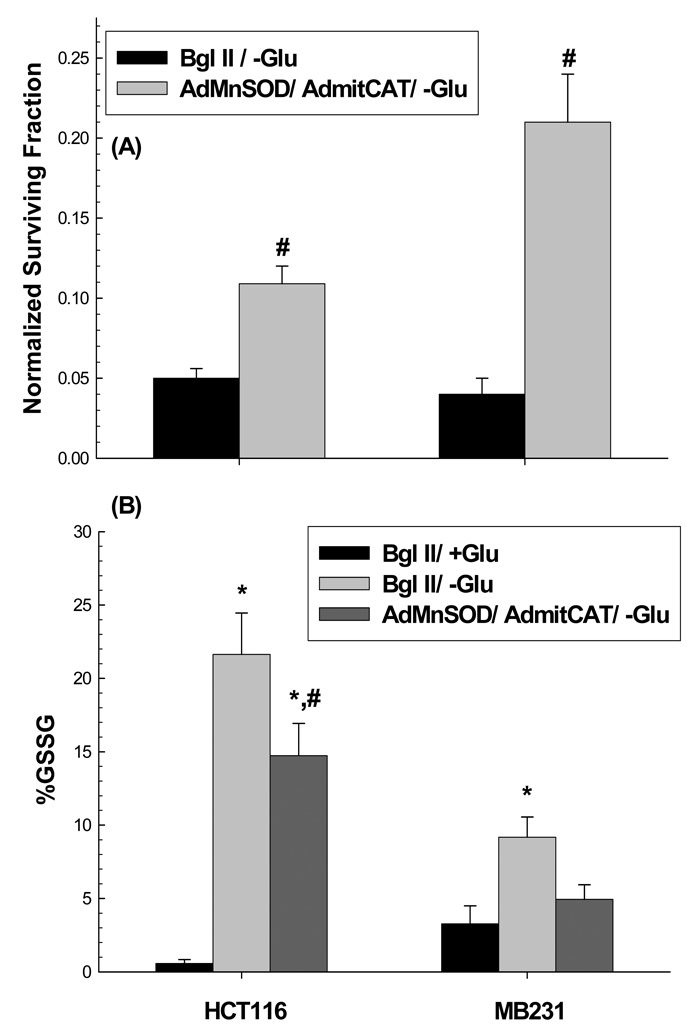
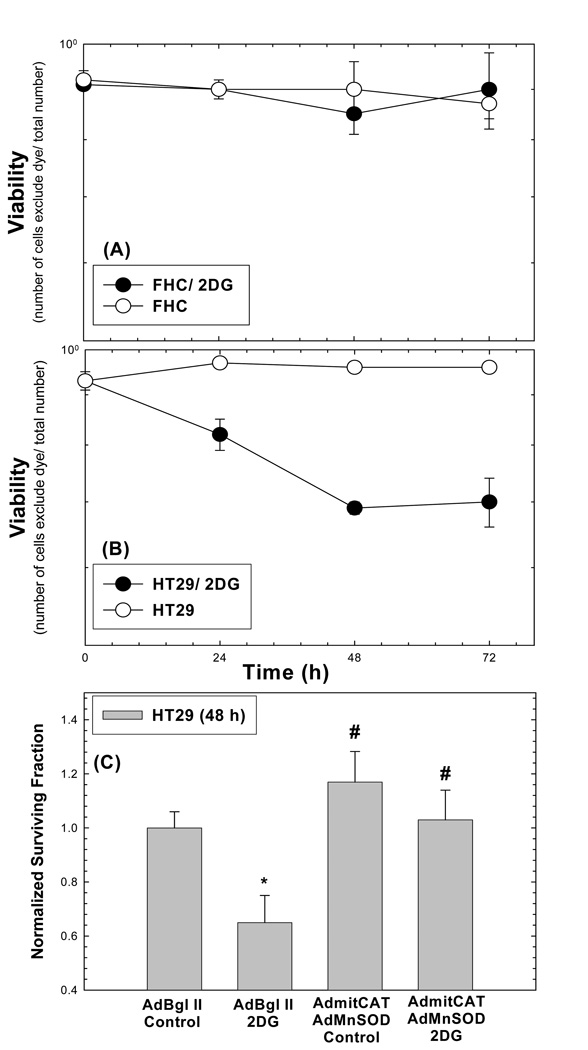
Cancer cells, relative to normal cells, demonstrate increased sensitivity to glucose-deprivation-induced cytotoxicity. To determine whether oxidative stress mediated by O(2)(*-) and hydroperoxides contributed to the differential susceptibility of human epithelial cancer cells to glucose deprivation, the oxidation of DHE (dihydroethidine; for O(2)(*-)) and CDCFH(2) [5- (and 6-)carboxy-2',7'-dichlorodihydrofluorescein diacetate; for hydroperoxides] was measured in human colon and breast cancer cells (HT29, HCT116, SW480 and MB231) and compared with that in normal human cells [FHC cells, 33Co cells and HMECs (human mammary epithelial cells)]. Cancer cells showed significant increases in DHE (2-20-fold) and CDCFH(2) (1.8-10-fold) oxidation, relative to normal cells, that were more pronounced in the presence of the mitochondrial electron-transport-chain blocker, antimycin A. Furthermore, HCT116 and MB231 cells were more susceptible to glucose-deprivation-induced cytotoxicity and oxidative stress, relative to 33Co cells and HMECs. HT29 cells were also more susceptible to 2DG (2-deoxyglucose)-induced cytotoxicity, relative to FHC cells. Overexpression of manganese SOD (superoxide dismutase) and mitochondrially targeted catalase significantly protected HCT116 and MB231 cells from glucose-deprivation-induced cytotoxicity and oxidative stress and also protected HT29 cells from 2DG-induced cytotoxicity. These results show that cancer cells (relative to normal cells) demonstrate increased steady-state levels of ROS (reactive oxygen species; i.e. O(2)(*-) and H(2)O(2)) that contribute to differential susceptibility to glucose-deprivation-induced cytotoxicity and oxidative stress. These studies support the hypotheses that cancer cells increase glucose metabolism to compensate for excess metabolic production of ROS and that inhibition of glucose and hydroperoxide metabolism may provide a biochemical target for selectively enhancing cytotoxicity and oxidative stress in human cancer cells.
Figures
References
-
- Lehninger AL. Biochemistry. New York: Worth Publishers Inc.; 1976.
-
- Warburg O. On the origin of cancer cells. Science. 1956;132:309–314. - PubMed
-
- Weber G. Enzymology of cancer cells (first of two parts) New Engl. J. Med. 1977;296:486–492. - PubMed
-
- Weber G. Enzymology of cancer cells (second of two parts) New Engl. J. Med. 1977;296:541–551. - PubMed
-
- Boveris A, Cadenas E. Production of superoxide radicals and hydrogen peroxide in mitochondria (Chapter 2) In: Oberley LW, editor. Superoxide Dimsutase. Boca Raton: CRC Press; 1982. pp. 15–30.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical