

Decreased expression of CD200 and CD200 receptor in Alzheimer's disease: a potential mechanism leading to chronic inflammation
- PMID: 18938162
- PMCID: PMC2765462
- DOI: 10.1016/j.expneurol.2008.09.003
Decreased expression of CD200 and CD200 receptor in Alzheimer's disease: a potential mechanism leading to chronic inflammation
Abstract
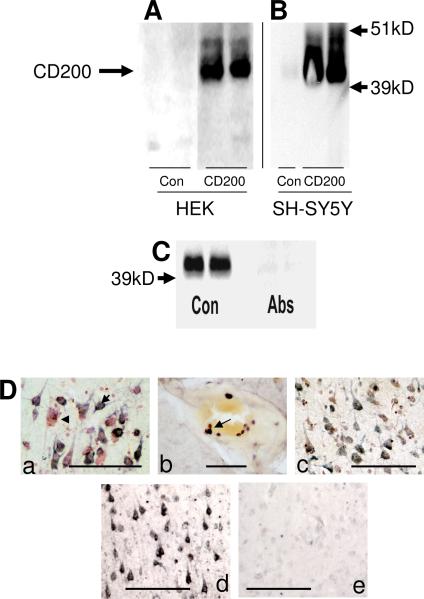
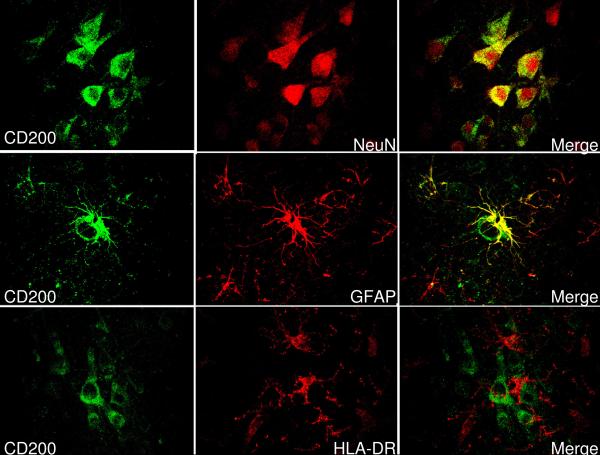
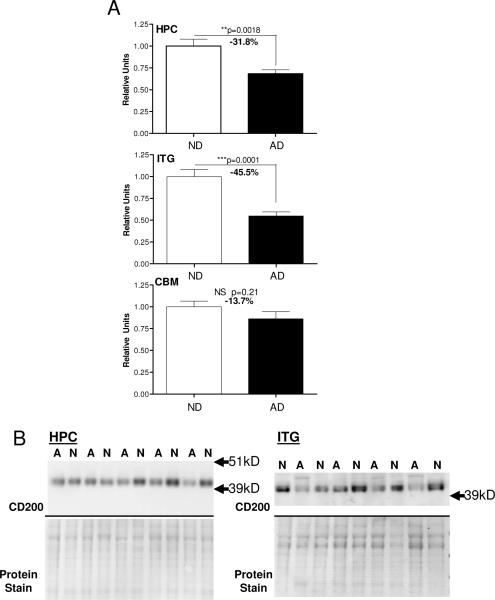
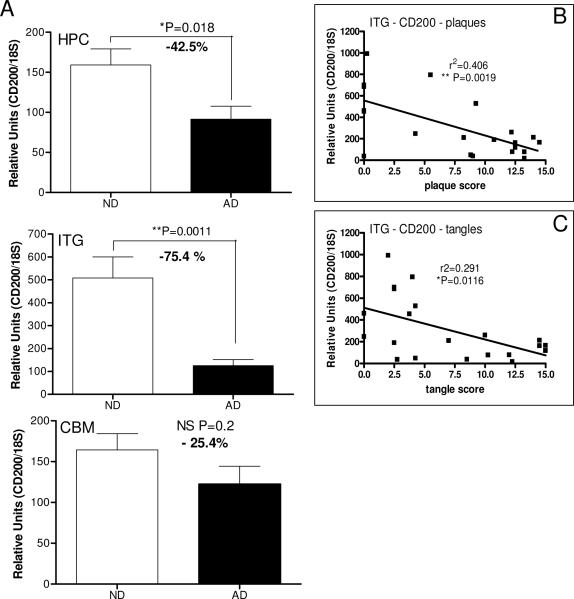
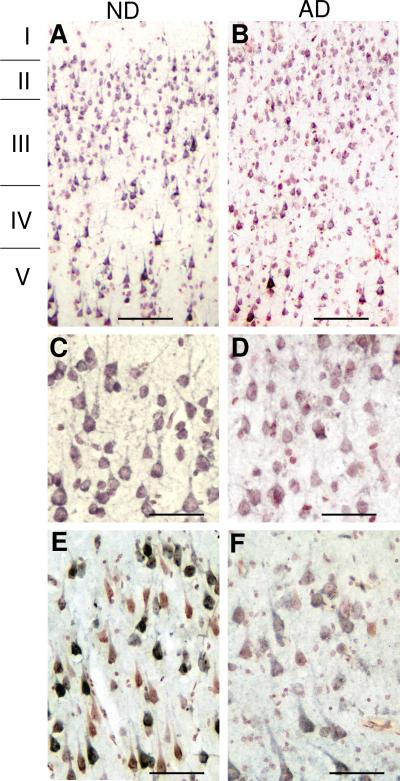
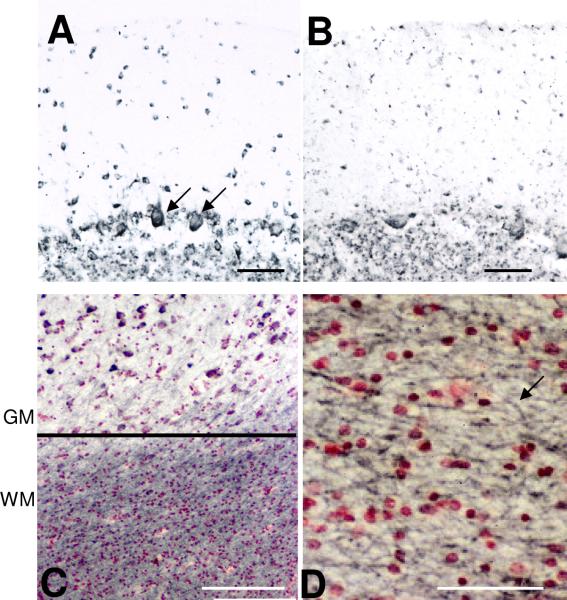
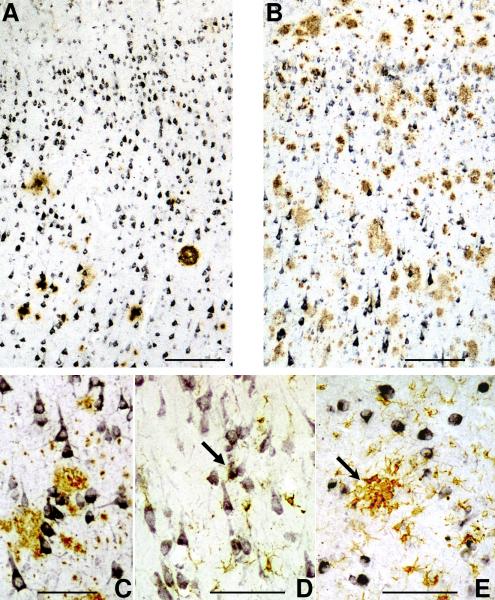
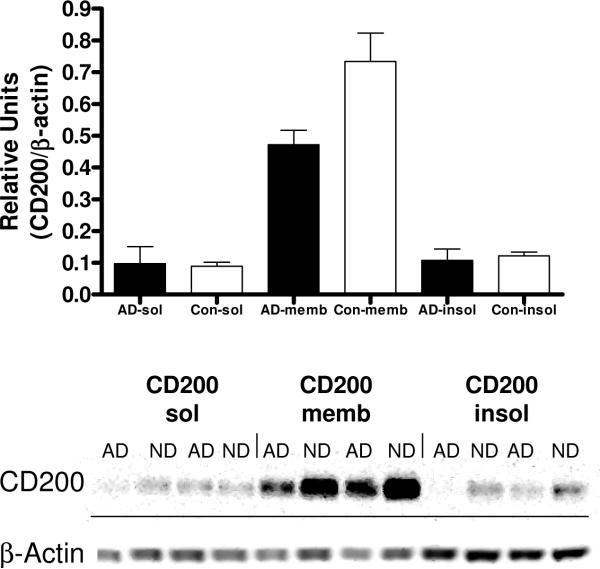
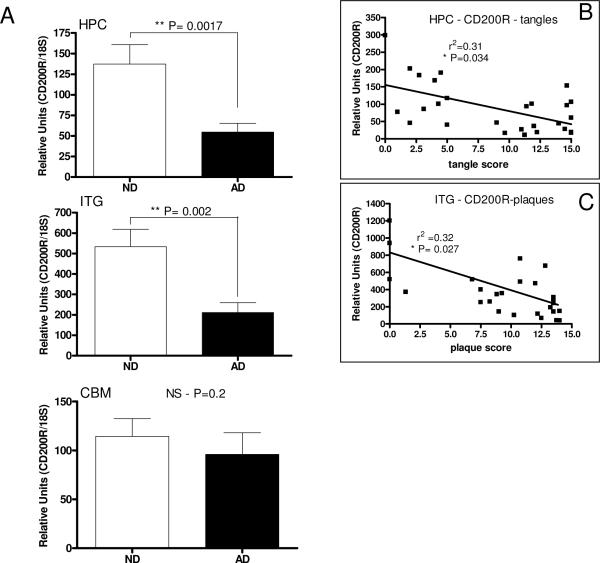
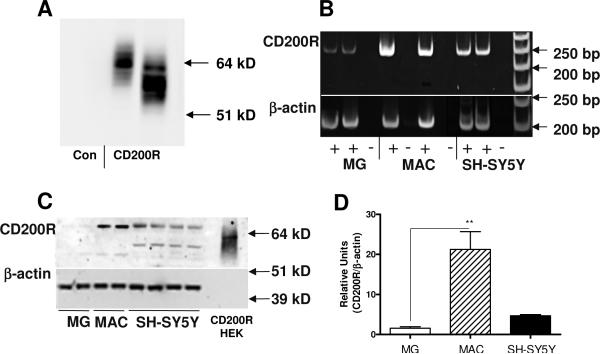
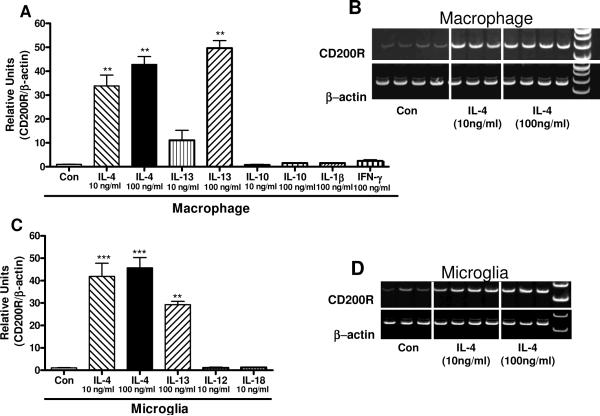
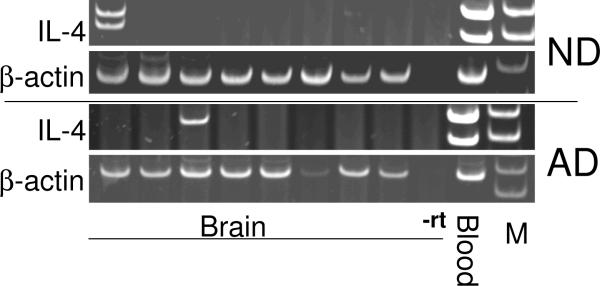
Inflammatory activation of microglia in response to neurodegenerative changes in diseases such as Alzheimer's disease (AD) and Parkinson's disease has been extensively described. These observations have suggested that inflammation could be contributing to disease progression. In this paper, the potential role of CD200 and CD200 receptor (CD200R), whose known functions are to activate anti-inflammatory pathways and induce immune tolerance through binding of CD200 to CD200 receptor (CD200R), was studied in AD. Quantitative studies showed a significant decrease in CD200 protein and mRNA in AD hippocampus and inferior temporal gyrus, but not cerebellum. Immunohistochemistry of brain tissue sections of hippocampus, superior frontal gyrus, inferior temporal gyrus and cerebellum from AD and non-demented cases demonstrated a predominant, though heterogeneous, neuronal localization for CD200. Decreased neuronal expression was apparent in brain regions affected by AD pathology. There was also a significant decrease in CD200R mRNA expression in AD hippocampus and inferior temporal gyrus, but not cerebellum. Low expression of CD200R by microglia was confirmed at the mRNA and protein level using cultured human microglia compared to blood-derived macrophages. Treatment of microglia and macrophages with interleukin-4 and interleukin-13 significantly increased expression of CD200R. Expression of these cytokines was not generally detectable in brain. These data indicate that the anti-inflammatory CD200/CD200R system may be deficient in AD brains. Mechanisms aimed at increasing levels of CD200 and CD200R could have therapeutic potential for controlling inflammation in human neurodegenerative diseases.
Figures
Similar articles
-
Distribution of the immune inhibitory molecules CD200 and CD200R in the normal central nervous system and multiple sclerosis lesions suggests neuron-glia and glia-glia interactions.J Neuropathol Exp Neurol. 2009 Feb;68(2):159-67. doi: 10.1097/NEN.0b013e3181964113. J Neuropathol Exp Neurol. 2009. PMID: 19151626
-
CD200-CD200R1 interaction contributes to neuroprotective effects of anandamide on experimentally induced inflammation.Glia. 2012 Sep;60(9):1437-50. doi: 10.1002/glia.22366. Epub 2012 May 31. Glia. 2012. PMID: 22653796
-
CD200-CD200R regulation of microglia activation in the pathogenesis of Parkinson's disease.J Neuroimmune Pharmacol. 2007 Sep;2(3):259-64. doi: 10.1007/s11481-007-9075-1. Epub 2007 May 18. J Neuroimmune Pharmacol. 2007. PMID: 18040859 Review.
-
Brain innate immunity in the regulation of neuroinflammation: therapeutic strategies by modulating CD200-CD200R interaction involve the cannabinoid system.Curr Pharm Des. 2014;20(29):4707-22. doi: 10.2174/1381612820666140130202911. Curr Pharm Des. 2014. PMID: 24588829 Free PMC article. Review.
-
Maternal Immune Activation Sensitizes Male Offspring Rats to Lipopolysaccharide-Induced Microglial Deficits Involving the Dysfunction of CD200-CD200R and CX3CL1-CX3CR1 Systems.Cells. 2020 Jul 12;9(7):1676. doi: 10.3390/cells9071676. Cells. 2020. PMID: 32664639 Free PMC article.
Cited by
-
The classification of microglial activation phenotypes on neurodegeneration and regeneration in Alzheimer's disease brain.Arch Immunol Ther Exp (Warsz). 2012 Aug;60(4):251-66. doi: 10.1007/s00005-012-0181-2. Epub 2012 Jun 19. Arch Immunol Ther Exp (Warsz). 2012. PMID: 22710659 Free PMC article. Review.
-
Targeting Impaired Antimicrobial Immunity in the Brain for the Treatment of Alzheimer's Disease.Neuropsychiatr Dis Treat. 2021 May 4;17:1311-1339. doi: 10.2147/NDT.S264910. eCollection 2021. Neuropsychiatr Dis Treat. 2021. PMID: 33976546 Free PMC article. Review.
-
Microglial Phenotyping in Neurodegenerative Disease Brains: Identification of Reactive Microglia with an Antibody to Variant of CD105/Endoglin.Cells. 2019 Jul 23;8(7):766. doi: 10.3390/cells8070766. Cells. 2019. PMID: 31340569 Free PMC article.
-
Altered expression of the immunoregulatory ligand-receptor pair CD200-CD200R1 in the brain of Parkinson's disease patients.NPJ Parkinsons Dis. 2022 Mar 16;8(1):27. doi: 10.1038/s41531-022-00290-2. NPJ Parkinsons Dis. 2022. PMID: 35296683 Free PMC article.
-
CD157 and CD200 at the crossroads of endothelial remodeling and immune regulation.Stem Cell Investig. 2019 Apr 19;6:10. doi: 10.21037/sci.2019.04.01. eCollection 2019. Stem Cell Investig. 2019. PMID: 31119148 Free PMC article. No abstract available.
References
-
- Banerjee D, Dick AD. Blocking CD200-CD200 receptor axis augments NOS-2 expression and aggravates experimental autoimmune uveoretinitis in Lewis rats. Ocul Immunol Inflamm. 2004;12:115–125. - PubMed
-
- Barclay AN, Wright GJ, Brooke G, Brown MH. CD200 and membrane protein interactions in the control of myeloid cells. Trends Immunol. 2002;23:285–290. - PubMed
-
- Boudakov I, Liu J, Fan N, Gulay P, Wong K, Gorczynski RM. Mice lacking CD200R1 show absence of suppression of lipopolysaccharide-induced tumor necrosis factor-alpha and mixed leukocyte culture responses by CD200. Transplantation. 2007;84:251–257. - PubMed
-
- Chen DX, Gorczynski RM. Discrete monoclonal antibodies define functionally important epitopes in the CD200 molecule responsible for immunosuppression function. Transplantation. 2005;79:282–288. - PubMed
-
- Chen J, Zhou Y, Mueller-Steiner S, Chen LF, Kwon H, Yi S, Mucke L, Gan L. SIRT1 protects against microglia-dependent amyloid-beta toxicity through inhibiting NF-kappaB signaling. J. Biol. Chem. 2005;280:40364–40374. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
