

Did RNA editing in plant organellar genomes originate under natural selection or through genetic drift?
- PMID: 18939975
- PMCID: PMC2584032
- DOI: 10.1186/1745-6150-3-43
Did RNA editing in plant organellar genomes originate under natural selection or through genetic drift?
Abstract
Background: The C<-->U substitution types of RNA editing have been observed frequently in organellar genomes of land plants. Although various attempts have been made to explain why such a seemingly inefficient genetic mechanism would have evolved, no satisfactory explanation exists in our view. In this study, we examined editing patterns in chloroplast genomes of the hornwort Anthoceros formosae and the fern Adiantum capillus-veneris and in mitochondrial genomes of the angiosperms Arabidopsis thaliana, Beta vulgaris and Oryza sativa, to gain an understanding of the question of how RNA editing originated.
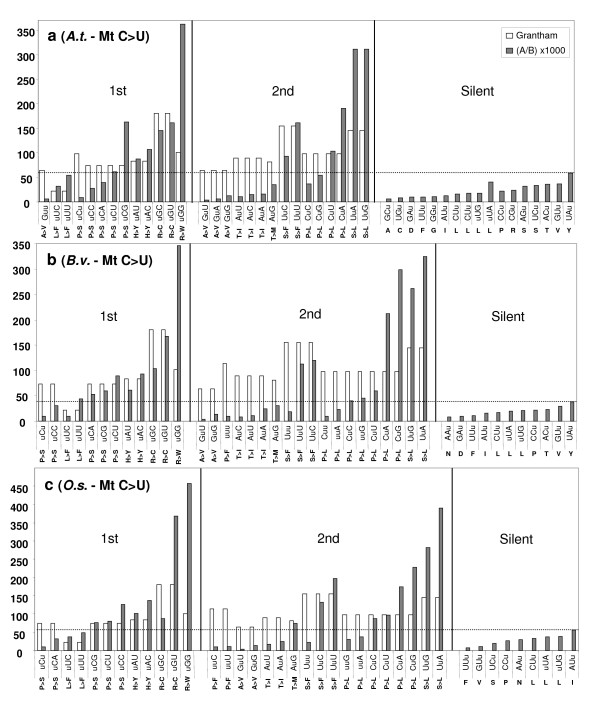
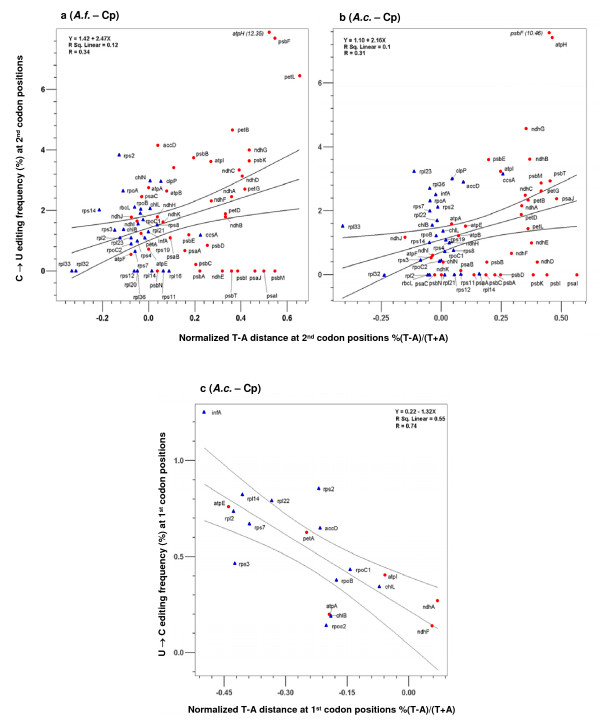
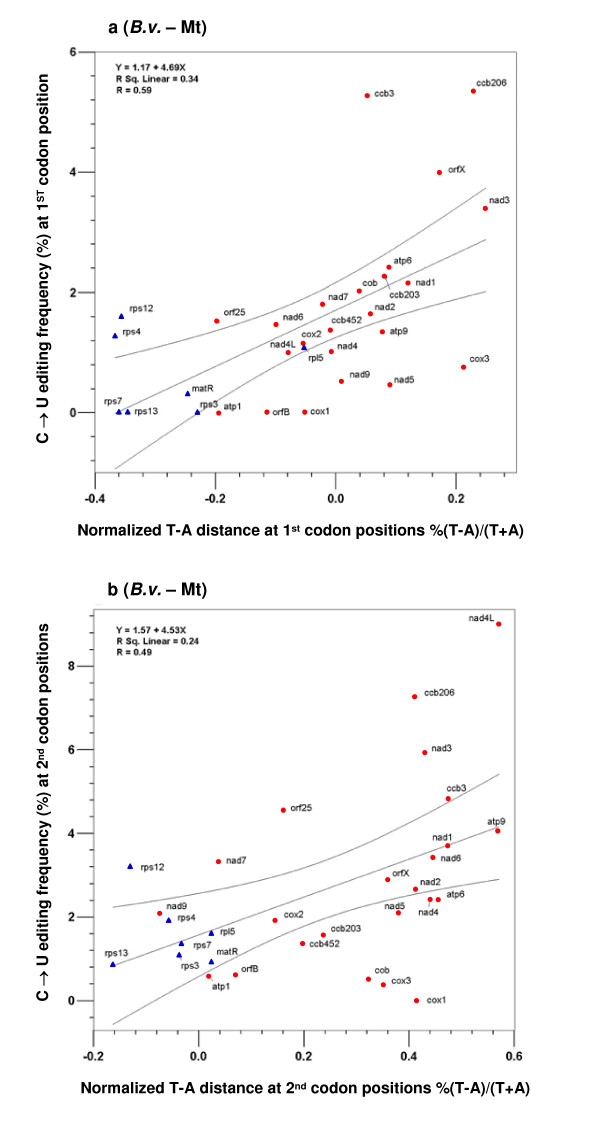
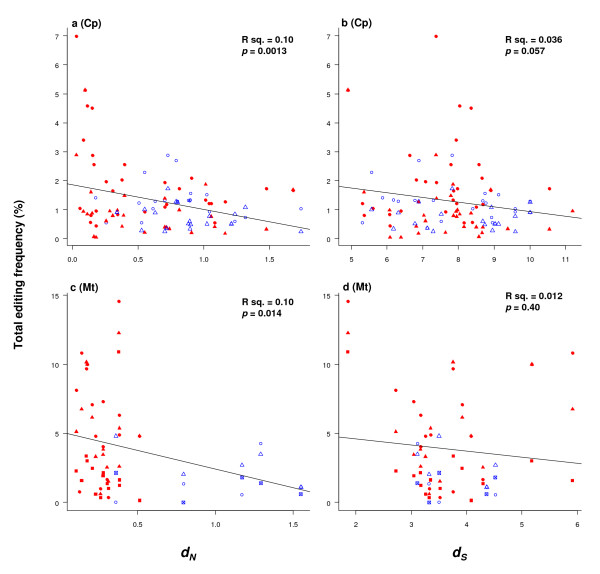
Results: We found that 1) most editing sites were distributed at the 2nd and 1st codon positions, 2) editing affected codons that resulted in larger hydrophobicity and molecular size changes much more frequently than those with little change involved, 3) editing uniformly increased protein hydrophobicity, 4) editing occurred more frequently in ancestrally T-rich sequences, which were more abundant in genes encoding membrane-bound proteins with many hydrophobic amino acids than in genes encoding soluble proteins, and 5) editing occurred most often in genes found to be under strong selective constraint.
Conclusion: These analyses show that editing mostly affects functionally important and evolutionarily conserved codon positions, codons and genes encoding membrane-bound proteins. In particular, abundance of RNA editing in plant organellar genomes may be associated with disproportionately large percentages of genes in these two genomes that encode membrane-bound proteins, which are rich in hydrophobic amino acids and selectively constrained. These data support a hypothesis that natural selection imposed by protein functional constraints has contributed to selective fixation of certain editing sites and maintenance of the editing activity in plant organelles over a period of more than four hundred millions years. The retention of genes encoding RNA editing activity may be driven by forces that shape nucleotide composition equilibrium in two organellar genomes of these plants. Nevertheless, the causes of lineage-specific occurrence of a large portion of RNA editing sites remain to be determined.
Figures


References
-
- Bass BL. RNA Editing: Frontiers in Molecular Biology. New York: Oxford University Press; 2001.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
