

V3 spinal neurons establish a robust and balanced locomotor rhythm during walking
- PMID: 18940590
- PMCID: PMC2753604
- DOI: 10.1016/j.neuron.2008.09.027
V3 spinal neurons establish a robust and balanced locomotor rhythm during walking
Abstract
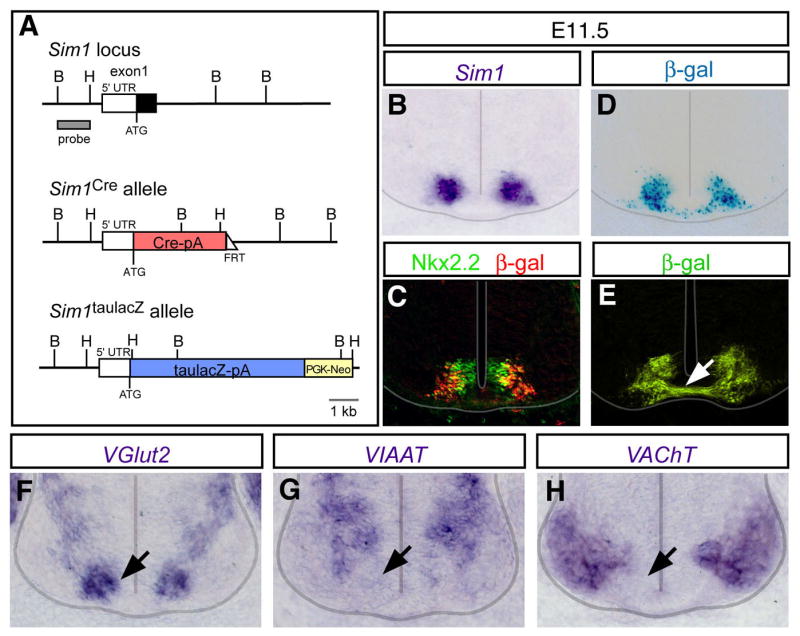
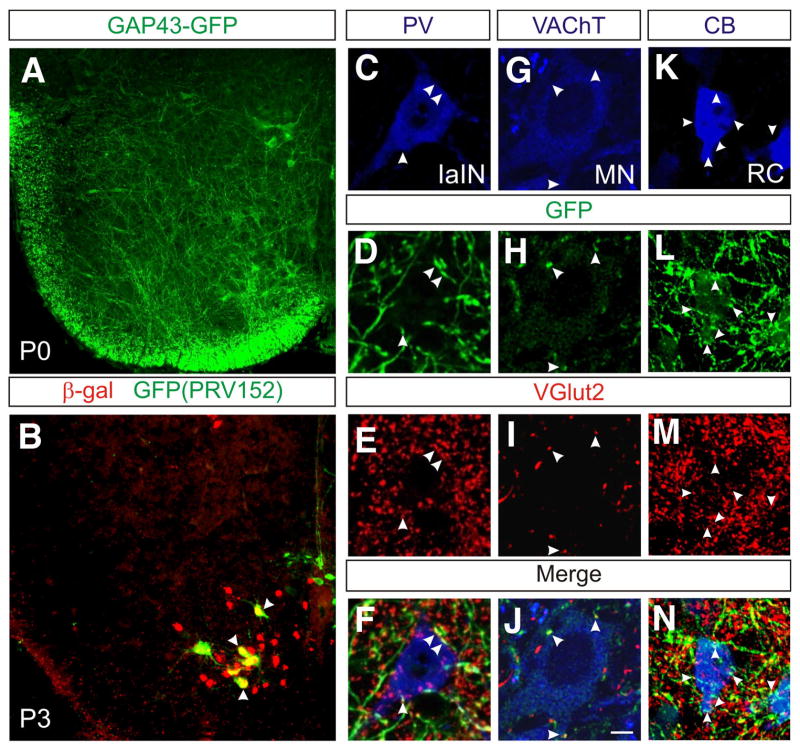
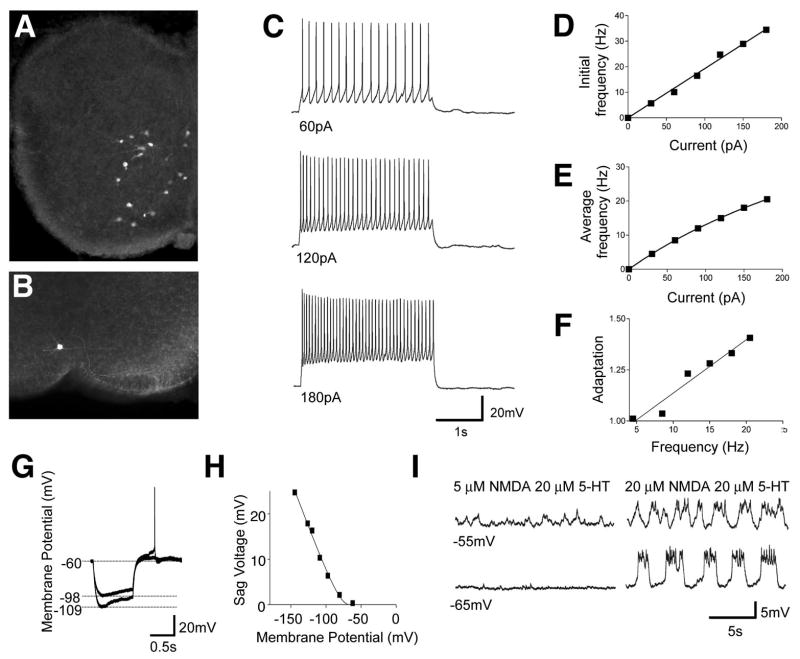
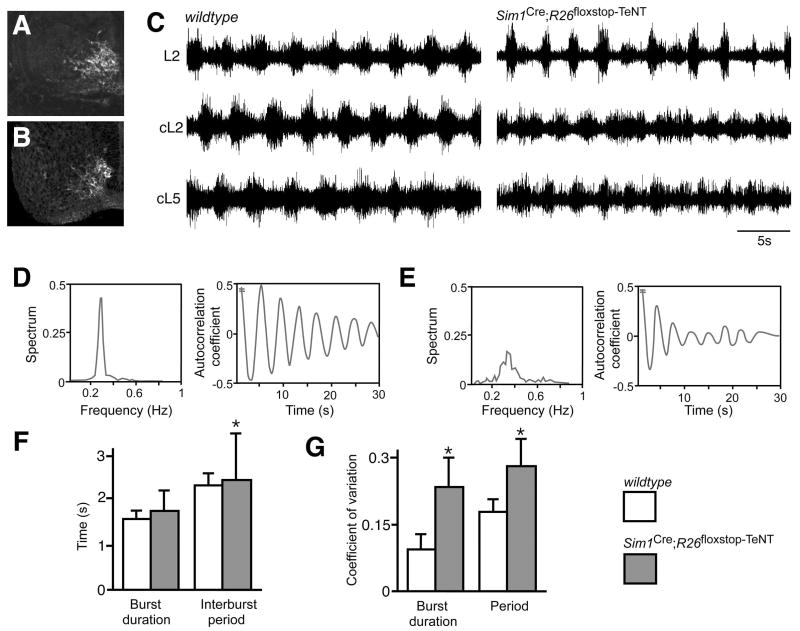
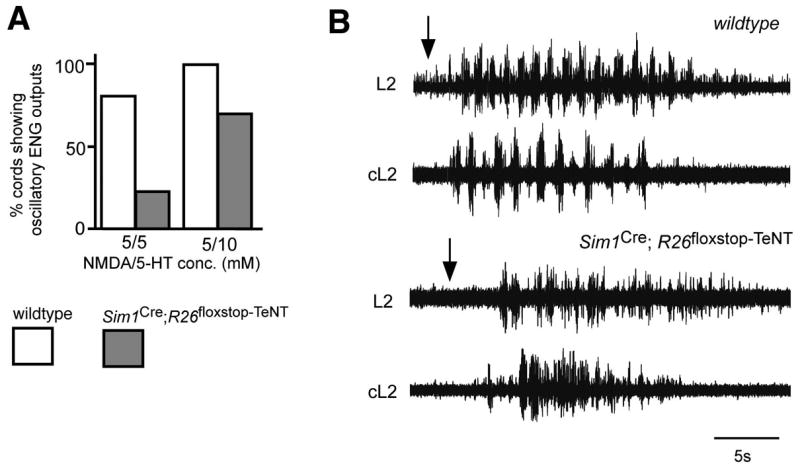
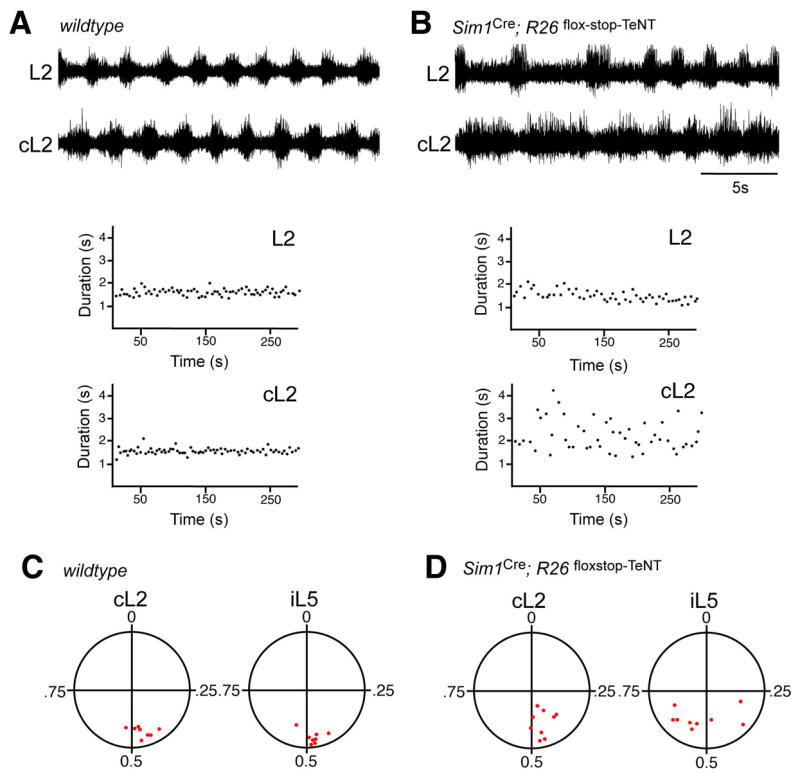
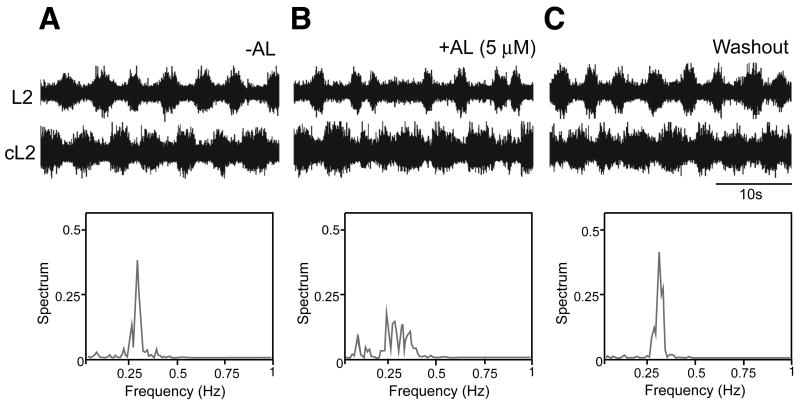
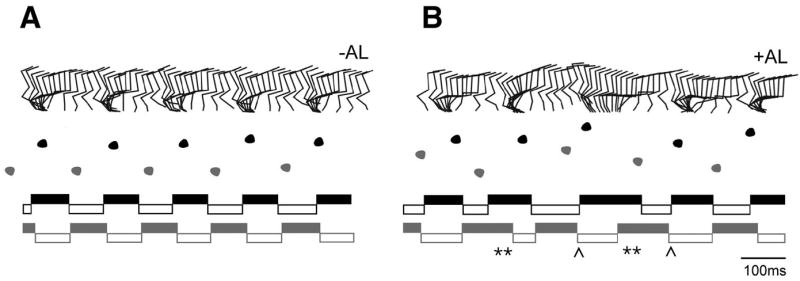
A robust and well-organized rhythm is a key feature of many neuronal networks, including those that regulate essential behaviors such as circadian rhythmogenesis, breathing, and locomotion. Here we show that excitatory V3-derived neurons are necessary for a robust and organized locomotor rhythm during walking. When V3-mediated neurotransmission is selectively blocked by the expression of the tetanus toxin light chain subunit (TeNT), the regularity and robustness of the locomotor rhythm is severely perturbed. A similar degeneration in the locomotor rhythm occurs when the excitability of V3-derived neurons is reduced acutely by ligand-induced activation of the allatostatin receptor. The V3-derived neurons additionally function to balance the locomotor output between both halves of the spinal cord, thereby ensuring a symmetrical pattern of locomotor activity during walking. We propose that the V3 neurons establish a regular and balanced motor rhythm by distributing excitatory drive between both halves of the spinal cord.
Figures
Comment in
-
Probing the locomotor conundrum: descending the 'V' interneuron ladder.Neuron. 2008 Oct 9;60(1):1-4. doi: 10.1016/j.neuron.2008.09.030. Neuron. 2008. PMID: 18940581 Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
