

An emerging role for voltage-gated Na+ channels in cellular migration: regulation of central nervous system development and potentiation of invasive cancers
- PMID: 18940784
- PMCID: PMC3380243
- DOI: 10.1177/1073858408320293
An emerging role for voltage-gated Na+ channels in cellular migration: regulation of central nervous system development and potentiation of invasive cancers
Abstract
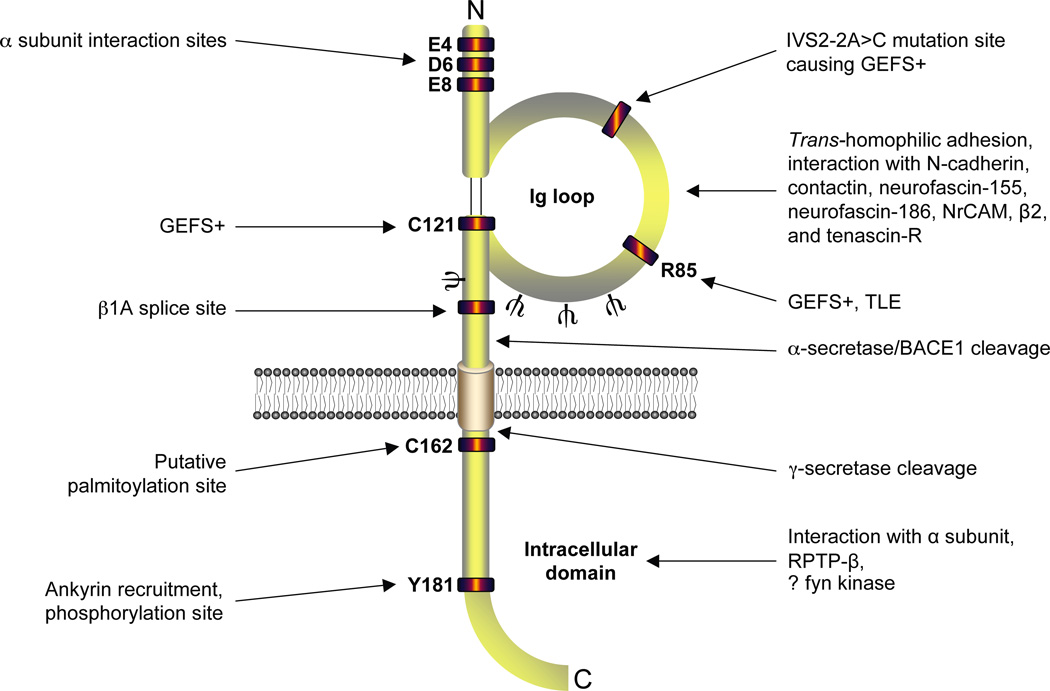
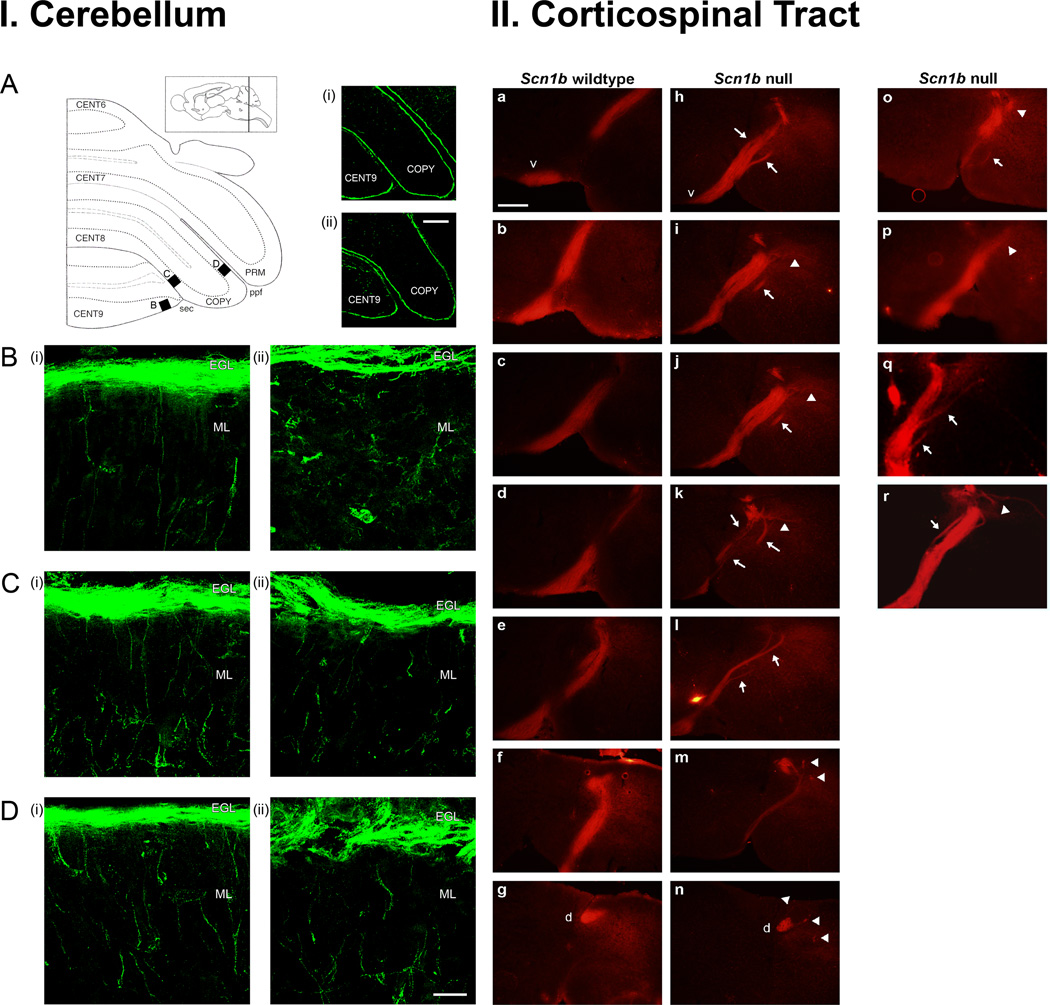
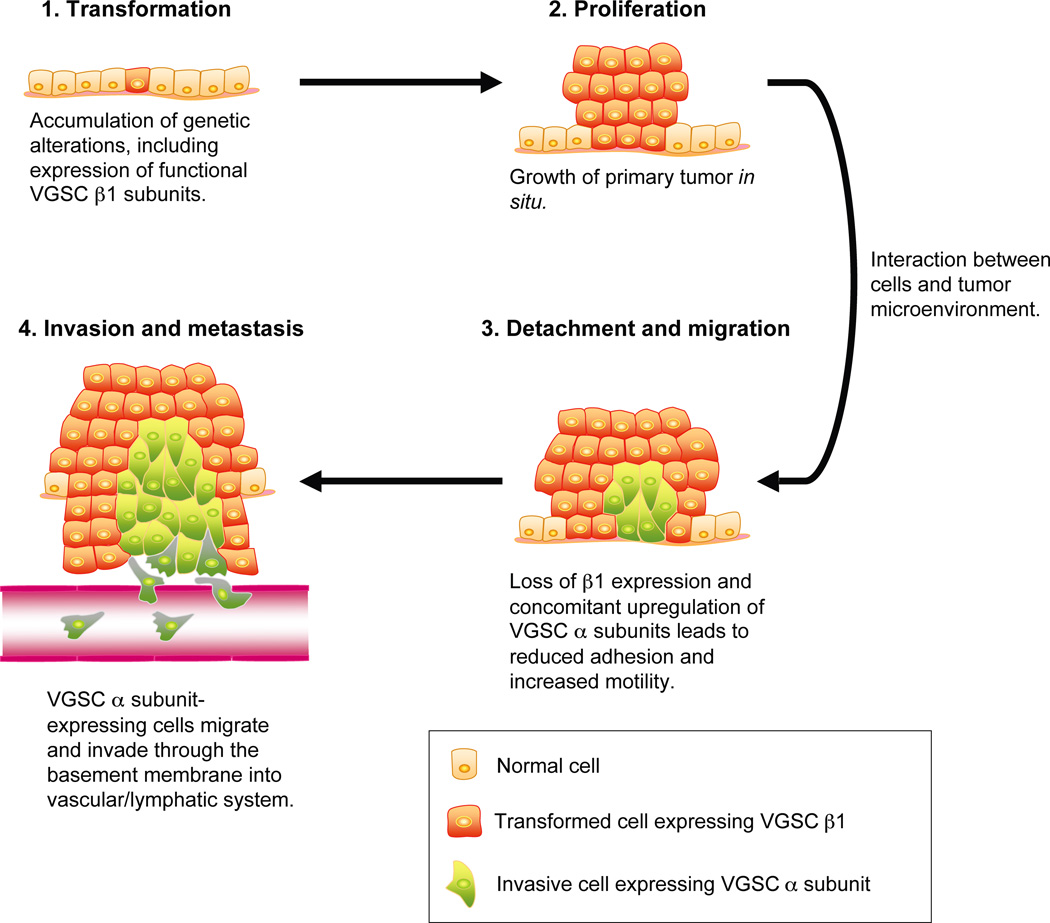
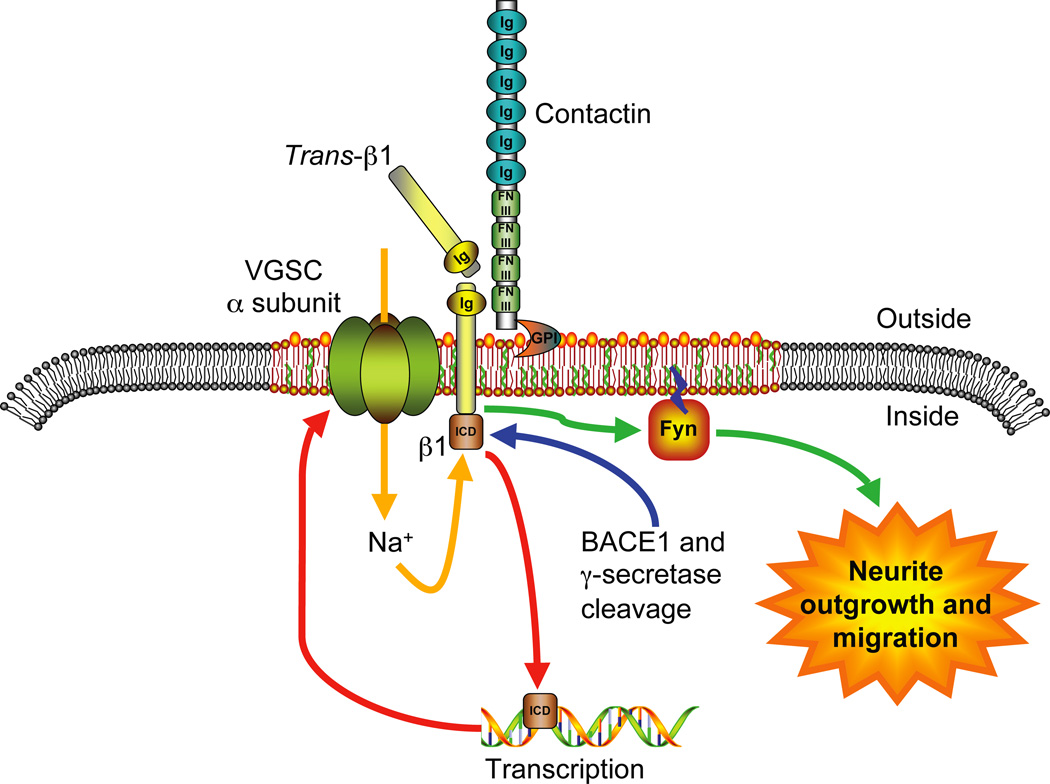
Voltage-gated Na(+) channels (VGSCs) exist as macromolecular complexes containing a pore-forming alpha subunit and one or more beta subunits. The VGSC alpha subunit gene family consists of 10 members, which have distinct tissue-specific and developmental expression profiles. So far, four beta subunits (beta1-beta4) and one splice variant of beta1 (beta1A, also called beta1B) have been identified. VGSC beta subunits are multifunctional, serving as modulators of channel activity, regulators of channel cell surface expression, and as members of the immunoglobulin superfamily, cell adhesion molecules (CAMs). beta subunits are substrates of beta-amyloid precursor protein-cleaving enzyme (BACE1) and gamma-secretase, yielding intracellular domains (ICDs) that may further modulate cellular activity via transcription. Recent evidence shows that beta1 regulates migration and pathfinding in the developing postnatal CNS in vivo. The alpha and beta subunits, together with other components of the VGSC signaling complex, may have dynamic interactive roles depending on cell/tissue type, developmental stage, and pathophysiology. In addition to excitable cells like nerve and muscle, VGSC alpha and beta subunits are functionally expressed in cells that are traditionally considered nonexcitable, including glia, vascular endothelial cells, and cancer cells. In particular, the alpha subunits are up-regulated in line with metastatic potential and are proposed to enhance cellular migration and invasion. In contrast to the alpha subunits, beta1 is more highly expressed in weakly metastatic cancer cells, and evidence suggests that its expression enhances cellular adhesion. Thus, novel roles are emerging for VGSC alpha and beta subunits in regulating migration during normal postnatal development of the CNS as well as during cancer metastasis.
Figures
References
-
- Abdul M, Hoosein N. Inhibition by anticonvulsants of prostate-specific antigen and interleukin-6 secretion by human prostate cancer cells. Anticancer Res. 2001;21(3B):2045–2048. - PubMed
-
- Abdul M, Hoosein N. Voltage-gated sodium ion channels in prostate cancer: expression and activity. Anticancer Res. 2002;22(3):1727–1730. - PubMed
-
- Abrahamsson PA. Neuroendocrine cells in tumour growth of the prostate. Endocr Relat Cancer. 1999;6(4):503–519. - PubMed
-
- Adachi K, Toyota M, Sasaki Y, Yamashita T, Ishida S, Ohe-Toyota M, et al. Identification of SCN3B as a novel p53-inducible proapoptotic gene. Oncogene. 2004;23(47):7791–7798. - PubMed
-
- Allen DH, Lepple-Wienhues A, Cahalan MD. Ion channel phenotype of melanoma cell lines. J Membr Biol. 1997;155(1):27–34. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
