

Evidence for core exosome independent function of the nuclear exoribonuclease Rrp6p
- PMID: 18940861
- PMCID: PMC2588529
- DOI: 10.1093/nar/gkn743
Evidence for core exosome independent function of the nuclear exoribonuclease Rrp6p
Abstract
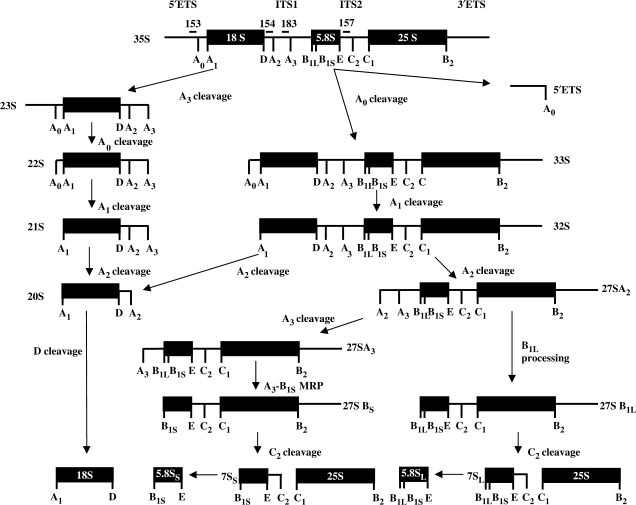
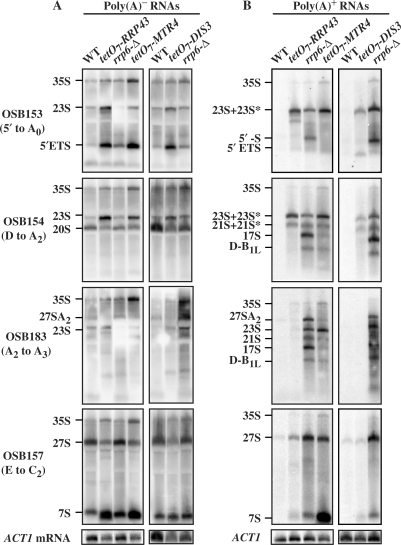
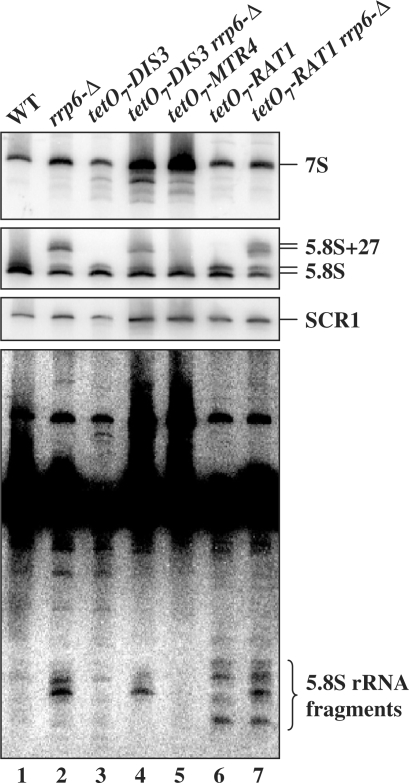
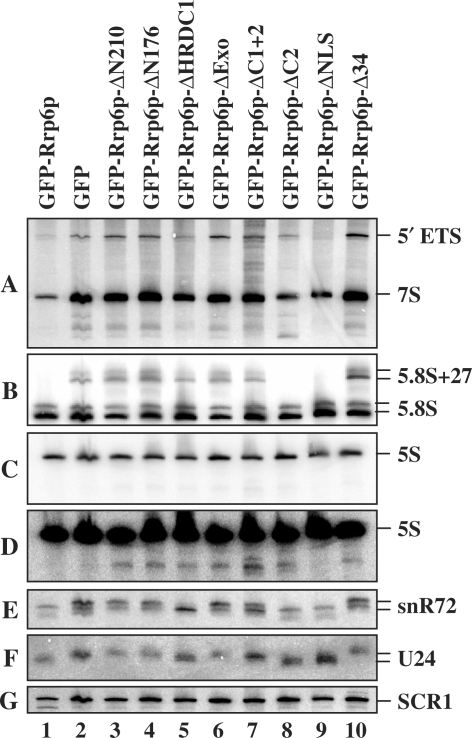
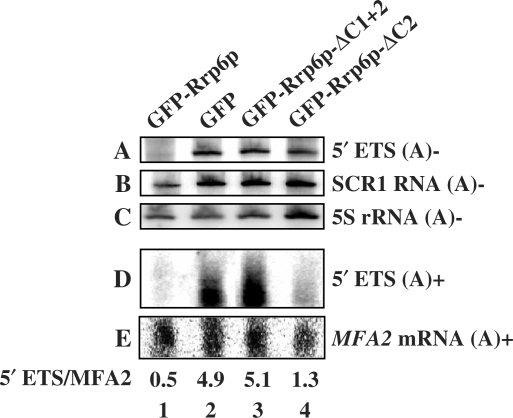
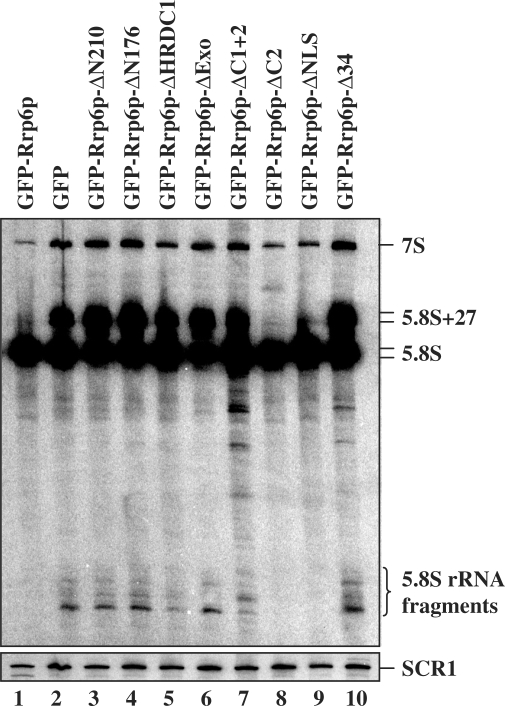
The RNA exosome processes and degrades RNAs in archaeal and eukaryotic cells. Exosomes from yeast and humans contain two active exoribonuclease components, Rrp6p and Dis3p/Rrp44p. Rrp6p is concentrated in the nucleus and the dependence of its function on the nine-subunit core exosome and Dis3p remains unclear. We found that cells lacking Rrp6p accumulate poly(A)+ rRNA degradation intermediates distinct from those found in cells depleted of Dis3p, or the core exosome component Rrp43p. Depletion of Dis3p in the absence of Rrp6p causes a synergistic increase in the levels of degradation substrates common to the core exosome and Rrp6p, but has no effect on Rrp6p-specific substrates. Rrp6p lacking a portion of its C-terminal domain no longer co-purifies with the core exosome, but continues to carry out RNA 3'-end processing of 5.8S rRNA and snoRNAs, as well as the degradation of certain truncated Rrp6-specific rRNA intermediates. However, disruption of Rrp6p-core exosome interaction results in the inability of the cell to efficiently degrade certain poly(A)+ rRNA processing products that require the combined activities of Dis3p and Rrp6p. These findings indicate that Rrp6p may carry out some of its critical functions without physical association with the core exosome.
Figures



References
-
- Moore MJ. Nuclear RNA turnover. Cell. 2002;108:431–434. - PubMed
-
- Amberg DC, Goldstein AL, Cole CN. Isolation and characterization of RAT1: an essential gene of Saccharomyces cerevisiae required for the efficient nucleocytoplasmic trafficking of mRNA. Genes Dev. 1992;6:1173–1189. - PubMed
-
- Kim M, Krogan NJ, Vasiljeva L, Rando OJ, Nedea E, Greenblatt JF, Buratowski S. The yeast Rat1 exonuclease promotes transcription termination by RNA polymerase II. Nature. 2004;432:517–522. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
