

Hepatitis C virus immune escape via exploitation of a hole in the T cell repertoire
- PMID: 18941234
- PMCID: PMC2742502
- DOI: 10.4049/jimmunol.181.9.6435
Hepatitis C virus immune escape via exploitation of a hole in the T cell repertoire
Abstract
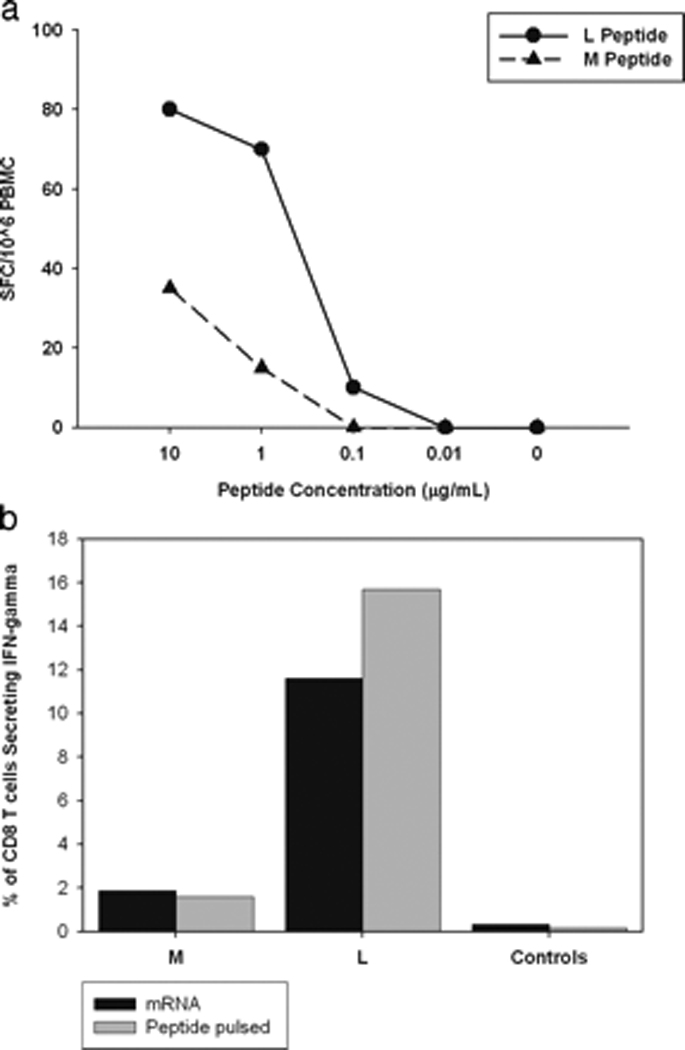
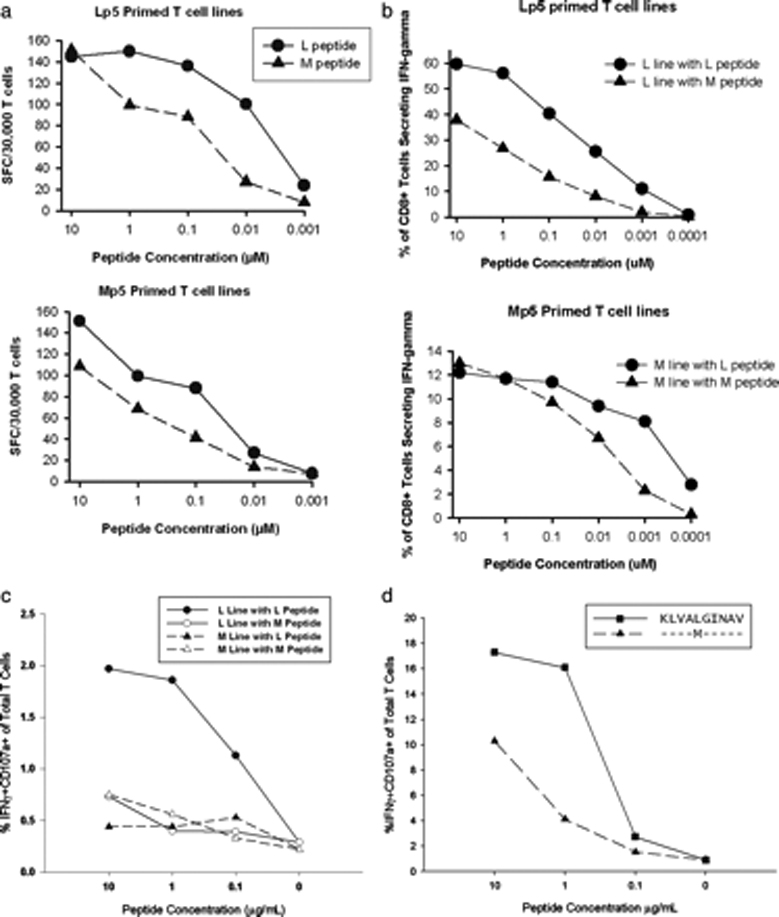
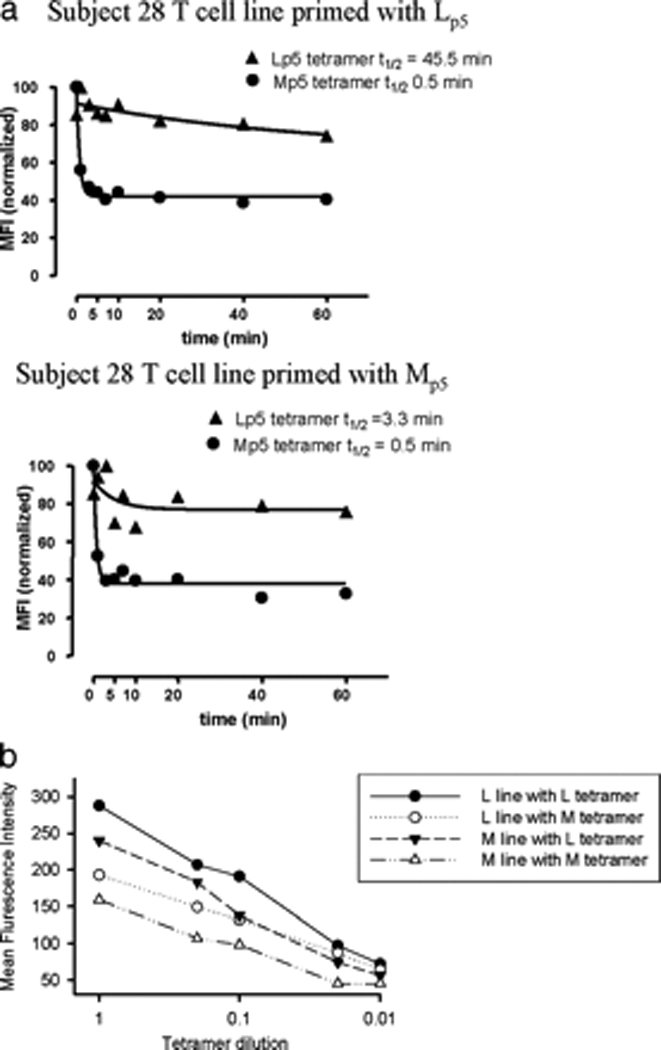
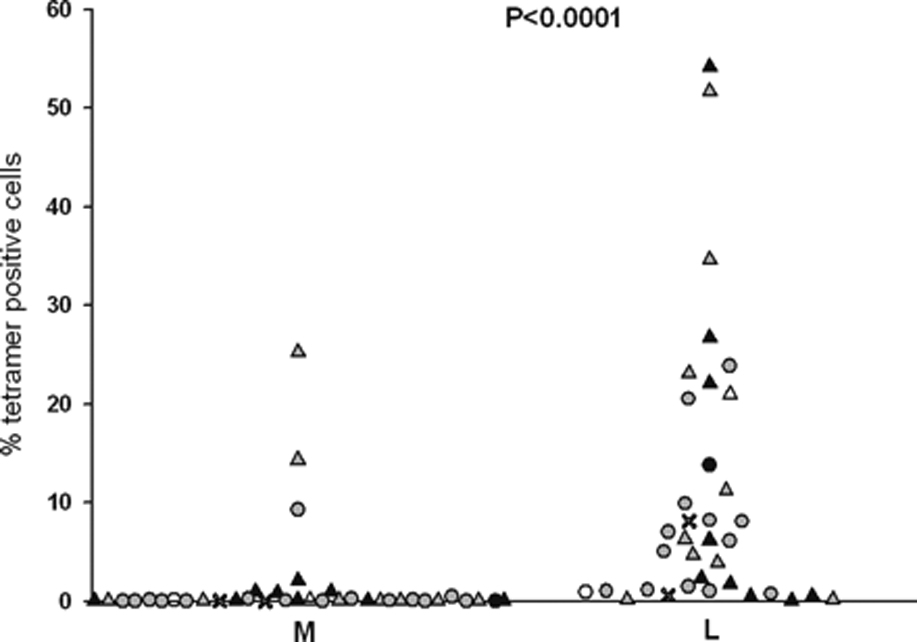
Hepatitis C virus (HCV) infection frequently persists despite eliciting substantial virus-specific immune responses. Thus, HCV infection provides a setting in which to investigate mechanisms of immune escape that allow for viral persistence. Viral amino acid substitutions resulting in decreased MHC binding or impaired Ag processing of T cell epitopes reduce Ag density on the cell surface, permitting evasion of T cell responses in chronic viral infection. Substitutions in viral epitopes that alter TCR contact residues frequently result in escape, but via unclear mechanisms because such substitutions do not reduce surface presentation of peptide-MHC complexes and would be expected to prime T cells with new specificities. We demonstrate that a known in vivo HCV mutation involving a TCR contact residue significantly diminishes T cell recognition and, in contrast to the original sequence, fails to effectively prime naive T cells. This mutant epitope thus escapes de novo immune recognition because there are few highly specific cognate TCR among the primary human T cell repertoire. This example is the first on viral immune escape via exploitation of a "hole" in the T cell repertoire, and may represent an important general mechanism of viral persistence.
Figures
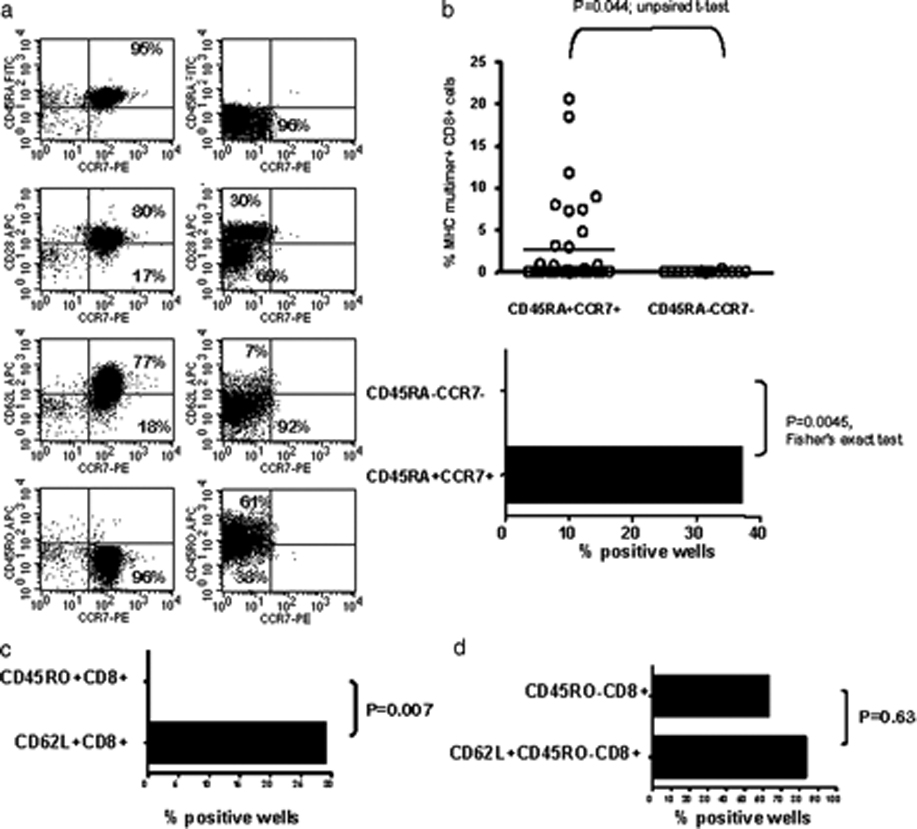
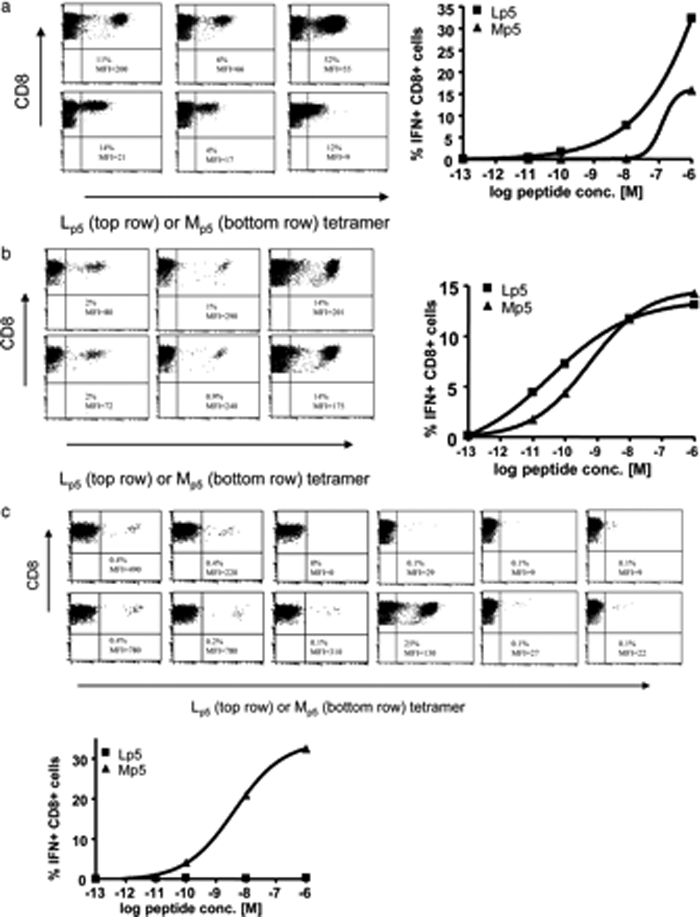
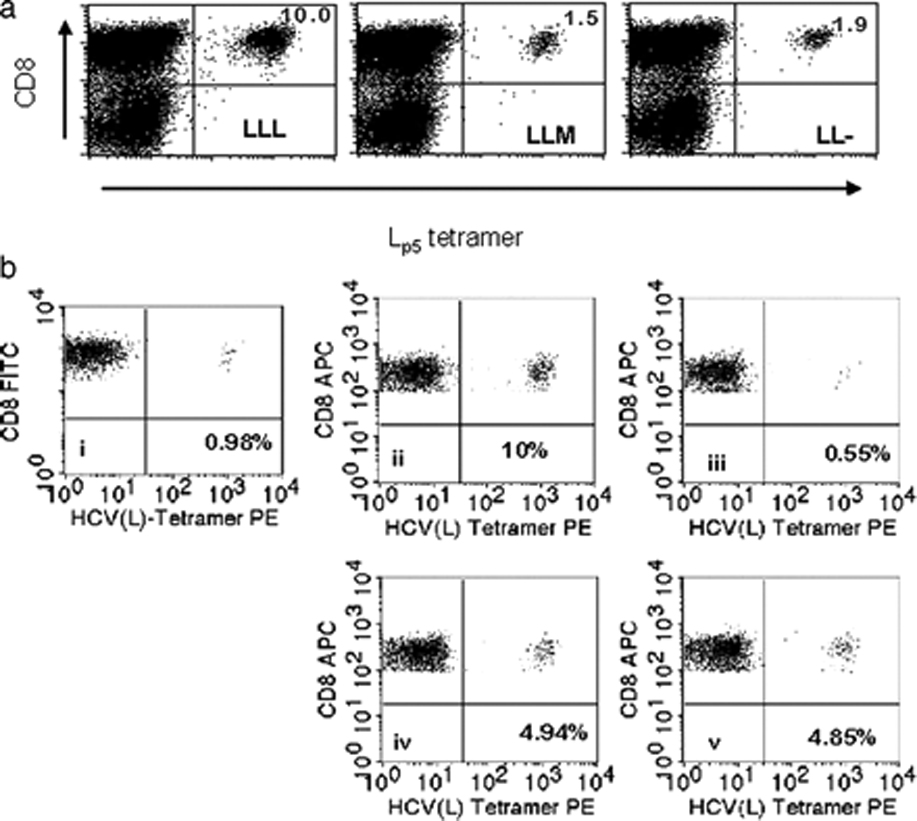
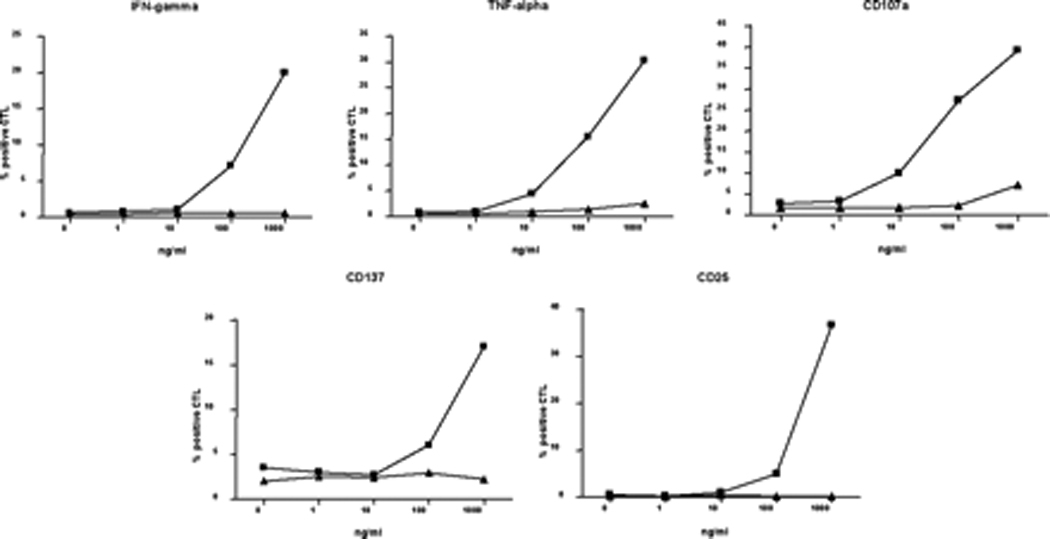
References
-
- Lechner F, Gruener NH, Urbani S, Uggeri J, Santantonio T, Kammer AR, Cerny A, Phillips R, Ferrari C, Pape GR, Klenerman P. CD8+ T lymphocyte responses are induced during acute hepatitis C virus infection but are not sustained. Eur.J.Immunol. 2000;30:2479–2487. - PubMed
-
- Cucchiarini M, Kammer AR, Grabscheid B, Diepolder HM, Gerlach TJ, Gruner N, Santantonio T, Reichen J, Pape GR, Cerny A. Vigorous peripheral blood cytotoxic T cell response during the acute phase of hepatitis C virus infection. Cell Immunol. 2000;203:111–123. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
