

Capture and transfer of HIV-1 particles by mature dendritic cells converges with the exosome-dissemination pathway
- PMID: 18945959
- PMCID: PMC2661860
- DOI: 10.1182/blood-2008-05-158642
Capture and transfer of HIV-1 particles by mature dendritic cells converges with the exosome-dissemination pathway
Abstract
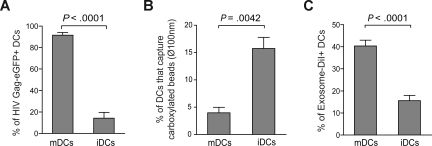
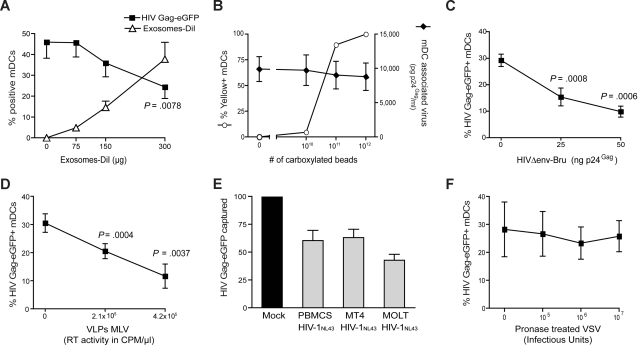
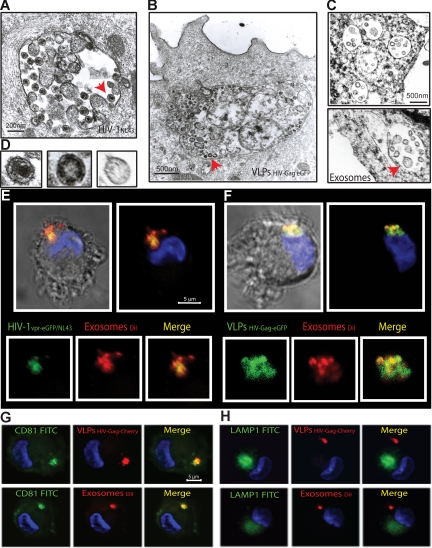
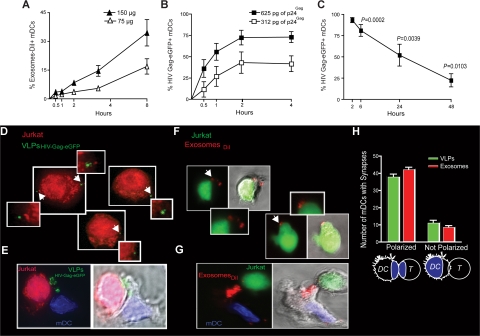
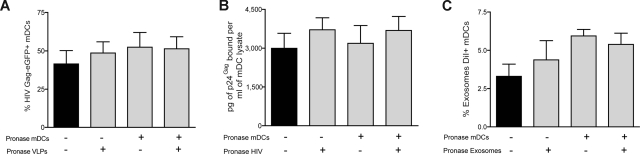
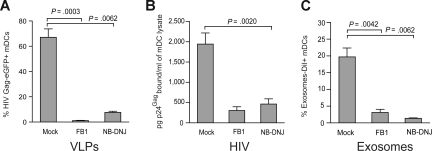
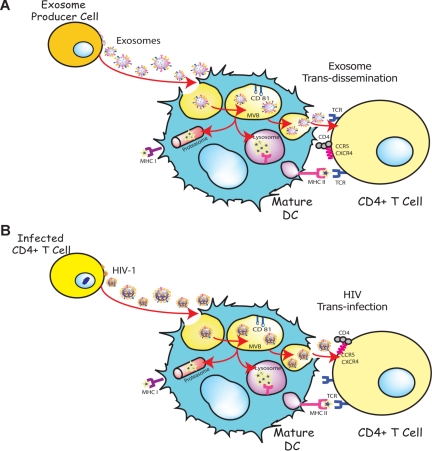
Exosomes are secreted cellular vesicles that can be internalized by dendritic cells (DCs), contributing to antigen-specific naive CD4(+) T-cell activation. Here, we demonstrate that human immunodeficiency virus type 1 (HIV-1) can exploit this exosome antigen-dissemination pathway intrinsic to mature DCs (mDCs) for mediating trans-infection of T lymphocytes. Capture of HIV-1, HIV-1 Gag-enhanced green fluorescent protein (eGFP) viral-like particles (VLPs), and exosomes by DCs was up-regulated upon maturation, resulting in localization within a CD81(+) compartment. Uptake of VLPs or exosomes could be inhibited by a challenge with either particle, suggesting that the expression of common determinant(s) on VLP or exosome surface is necessary for internalization by mDCs. Capture by mDCs was insensitive to proteolysis but blocked when virus, VLPs, or exosomes were produced from cells treated with sphingolipid biosynthesis inhibitors that modulate the lipid composition of the budding particles. Finally, VLPs and exosomes captured by mDCs were transmitted to T lymphocytes in an envelope glycoprotein-independent manner, underscoring a new potential viral dissemination pathway.
Figures
References
-
- Cameron PU, Freudenthal PS, Barker JM, Gezelter S, Inaba K, Steinman RM. Dendritic cells exposed to human immunodeficiency virus type-1 transmit a vigorous cytopathic infection to CD4+ T cells. Science. 1992;257:383–387. - PubMed
-
- Geijtenbeek TB, Kwon DS, Torensma R, et al. DC-SIGN, a dendritic cell-specific HIV-1-binding protein that enhances trans-infection of T cells. Cell. 2000;100:587–597. - PubMed
-
- McDonald D, Wu L, Bohks SM, Kewal Ramani VN, Unutmaz D, Hope TJ. Recruitment of HIV and its receptors to dendritic cell-T cell junctions. Science. 2003;300:1295–1297. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
