

Transcription inactivation through local refolding of the RNA polymerase structure
- PMID: 18946472
- PMCID: PMC2628454
- DOI: 10.1038/nature07510
Transcription inactivation through local refolding of the RNA polymerase structure
Abstract
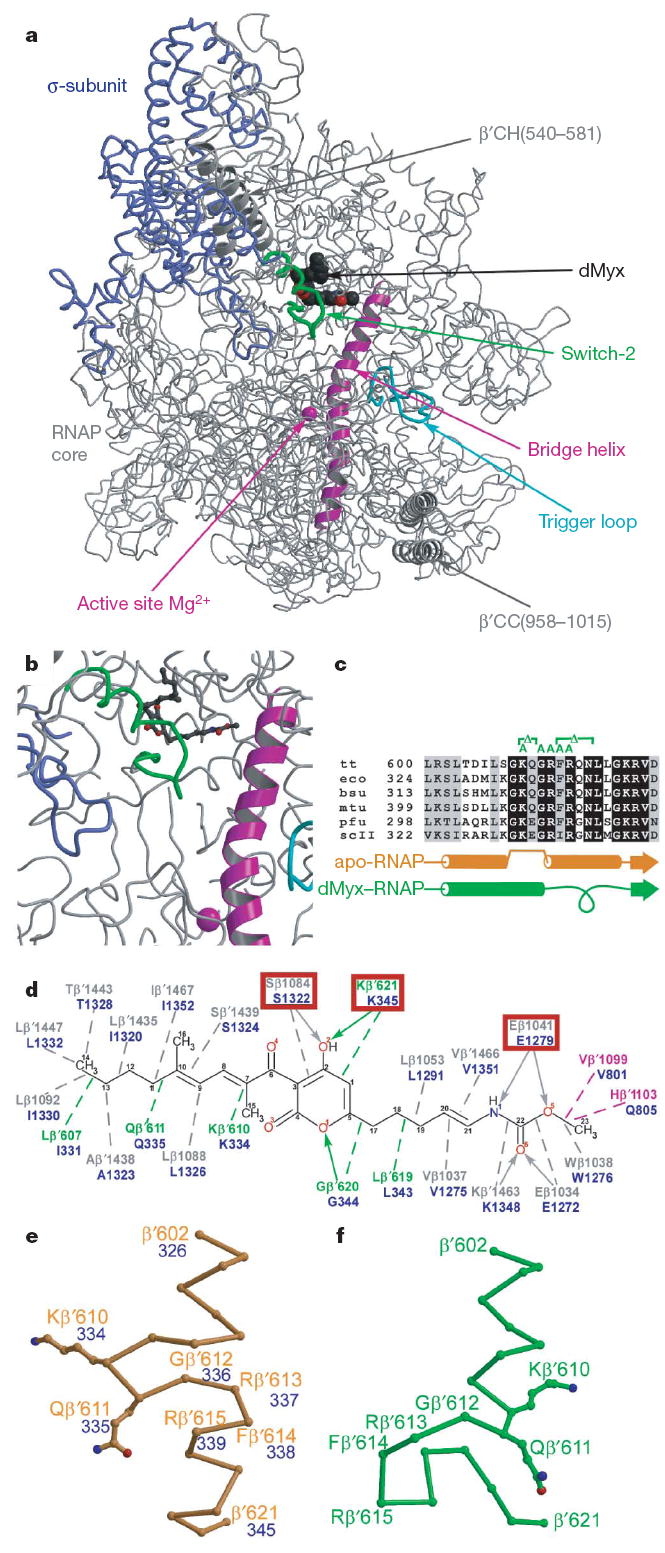
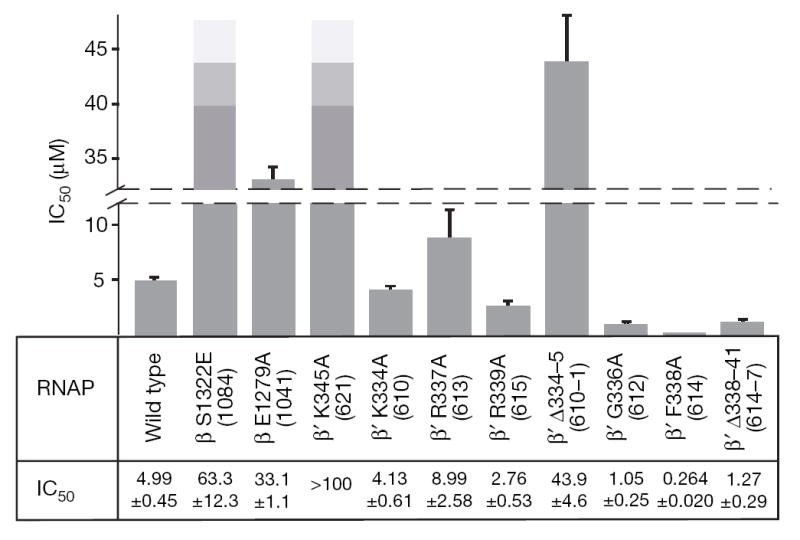
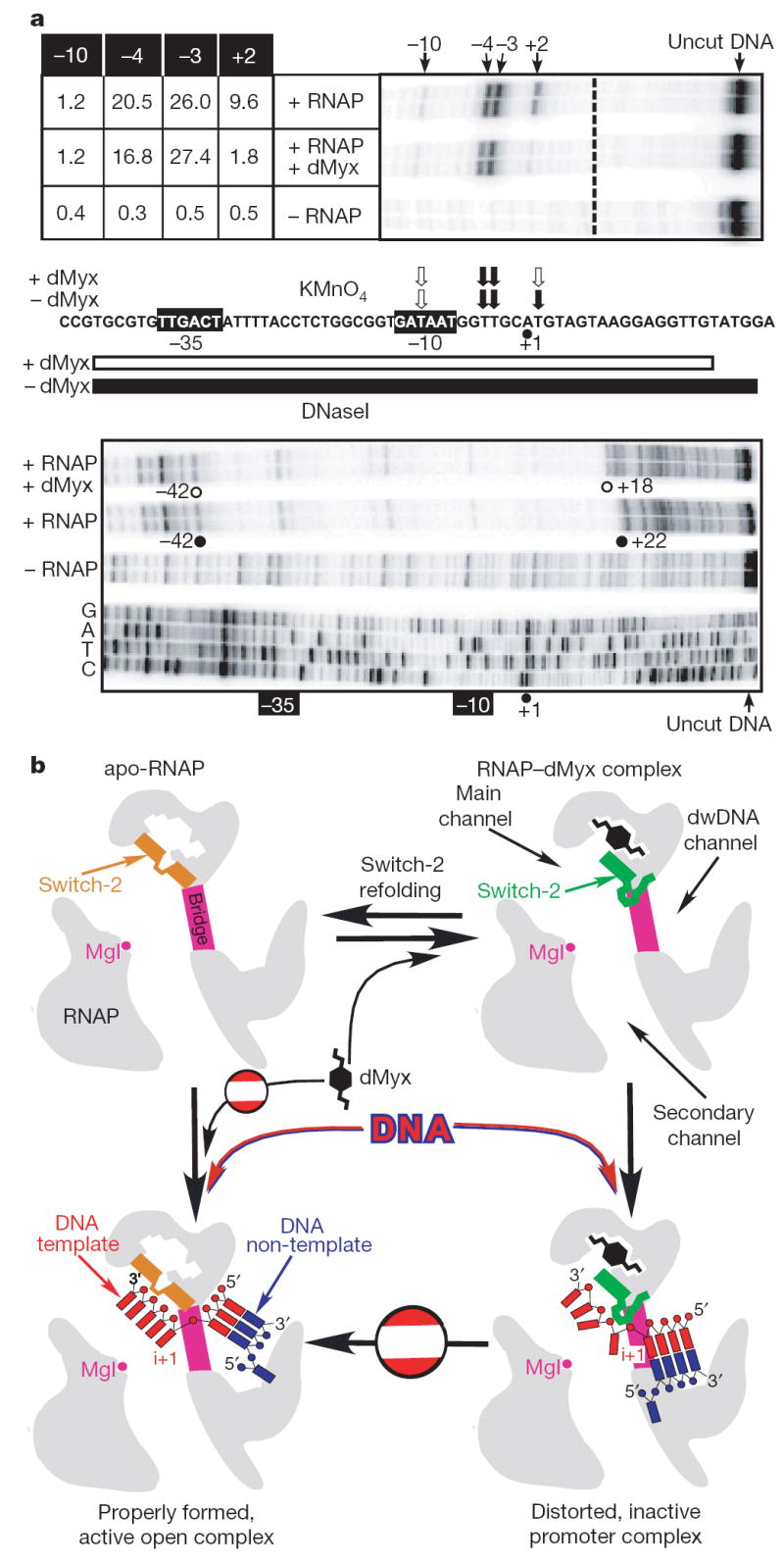
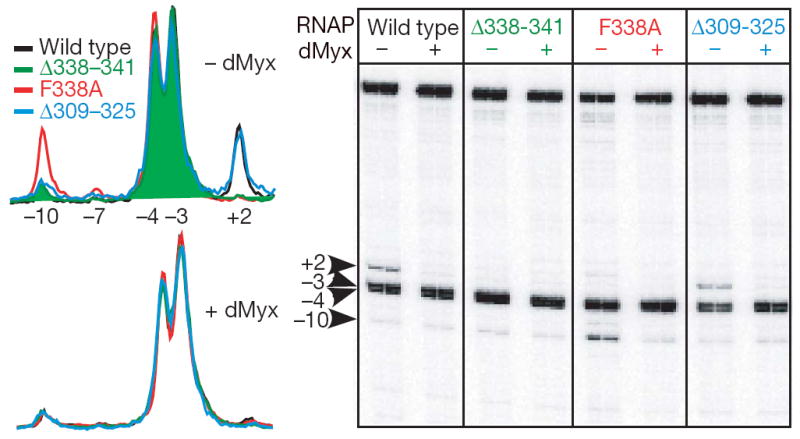
Structural studies of antibiotics not only provide a shortcut to medicine allowing for rational structure-based drug design, but may also capture snapshots of dynamic intermediates that become 'frozen' after inhibitor binding. Myxopyronin inhibits bacterial RNA polymerase (RNAP) by an unknown mechanism. Here we report the structure of dMyx--a desmethyl derivative of myxopyronin B--complexed with a Thermus thermophilus RNAP holoenzyme. The antibiotic binds to a pocket deep inside the RNAP clamp head domain, which interacts with the DNA template in the transcription bubble. Notably, binding of dMyx stabilizes refolding of the beta'-subunit switch-2 segment, resulting in a configuration that might indirectly compromise binding to, or directly clash with, the melted template DNA strand. Consistently, footprinting data show that the antibiotic binding does not prevent nucleation of the promoter DNA melting but instead blocks its propagation towards the active site. Myxopyronins are thus, to our knowledge, a first structurally characterized class of antibiotics that target formation of the pre-catalytic transcription initiation complex-the decisive step in gene expression control. Notably, mutations designed in switch-2 mimic the dMyx effects on promoter complexes in the absence of antibiotic. Overall, our results indicate a plausible mechanism of the dMyx action and a stepwise pathway of open complex formation in which core enzyme mediates the final stage of DNA melting near the transcription start site, and that switch-2 might act as a molecular checkpoint for DNA loading in response to regulatory signals or antibiotics. The universally conserved switch-2 may have the same role in all multisubunit RNAPs.
Figures
Similar articles
-
Subunit specialization in AAA+ proteins and substrate unfolding during transcription complex remodeling.Proc Natl Acad Sci U S A. 2025 Apr 29;122(17):e2425868122. doi: 10.1073/pnas.2425868122. Epub 2025 Apr 24. Proc Natl Acad Sci U S A. 2025. PMID: 40273105 Free PMC article.
-
The Black Book of Psychotropic Dosing and Monitoring.Psychopharmacol Bull. 2024 Jul 8;54(3):8-59. Psychopharmacol Bull. 2024. PMID: 38993656 Free PMC article. Review.
-
Characterization of a transitionally occupied state and thermal unfolding of domain 1.1 of factor of RNA polymerase from Bacillus subtilis.Proteins. 2023 Sep;91(9):1276-1287. doi: 10.1002/prot.26531. Epub 2023 Jun 22. Proteins. 2023. PMID: 37350110
-
Sexual Harassment and Prevention Training.2024 Mar 29. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. 2024 Mar 29. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. PMID: 36508513 Free Books & Documents.
-
Systemic pharmacological treatments for chronic plaque psoriasis: a network meta-analysis.Cochrane Database Syst Rev. 2021 Apr 19;4(4):CD011535. doi: 10.1002/14651858.CD011535.pub4. Cochrane Database Syst Rev. 2021. Update in: Cochrane Database Syst Rev. 2022 May 23;5:CD011535. doi: 10.1002/14651858.CD011535.pub5. PMID: 33871055 Free PMC article. Updated.
Cited by
-
Functional analysis of Thermus thermophilus transcription factor NusG.Nucleic Acids Res. 2010 Nov;38(21):7432-45. doi: 10.1093/nar/gkq623. Epub 2010 Jul 17. Nucleic Acids Res. 2010. PMID: 20639538 Free PMC article.
-
New target for inhibition of bacterial RNA polymerase: 'switch region'.Curr Opin Microbiol. 2011 Oct;14(5):532-43. doi: 10.1016/j.mib.2011.07.030. Epub 2011 Aug 19. Curr Opin Microbiol. 2011. PMID: 21862392 Free PMC article. Review.
-
Structural Basis of Transcription Inhibition by Fidaxomicin (Lipiarmycin A3).Mol Cell. 2018 Apr 5;70(1):60-71.e15. doi: 10.1016/j.molcel.2018.02.026. Epub 2018 Mar 29. Mol Cell. 2018. PMID: 29606590 Free PMC article.
-
A Search for Dual Action HIV-1 Reverse Transcriptase, Bacterial RNA Polymerase Inhibitors.Molecules. 2017 Oct 25;22(11):1808. doi: 10.3390/molecules22111808. Molecules. 2017. PMID: 29068373 Free PMC article.
-
Targeting Bacterial RNA Polymerase: Harnessing Simulations and Machine Learning to Design Inhibitors for Drug-Resistant Pathogens.Biochemistry. 2025 Mar 18;64(6):1169-1179. doi: 10.1021/acs.biochem.4c00751. Epub 2025 Feb 27. Biochemistry. 2025. PMID: 40014017 Review.
References
-
- Brueckner F, Cramer P. Structural basis of transcription inhibition by α-amanitin and implications for RNA polymerase II translocation. Nature Struct Mol Biol. 2008;15:811–818. - PubMed
-
- Vassylyev DG, et al. Structural basis for substrate loading in bacterial RNA polymerase. Nature. 2007;448:163–168. - PubMed
-
- Irschik H, Gerth K, Hofle G, Kohl W, Reichenbach H. The myxopyronins, new inhibitors of bacterial RNA synthesis from Myxococcus fulvus (Myxobacterales) J Antibiot (Tokyo) 1983;36:1651–1658. - PubMed
-
- Lira R, et al. Syntheses of novel myxopyronin B analogs as potential inhibitors of bacterial RNA polymerase. Bioorg Med Chem Lett. 2007;17:6797–6800. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
