

Splicing factors facilitate RNAi-directed silencing in fission yeast
- PMID: 18948543
- PMCID: PMC2585287
- DOI: 10.1126/science.1164029
Splicing factors facilitate RNAi-directed silencing in fission yeast
Abstract
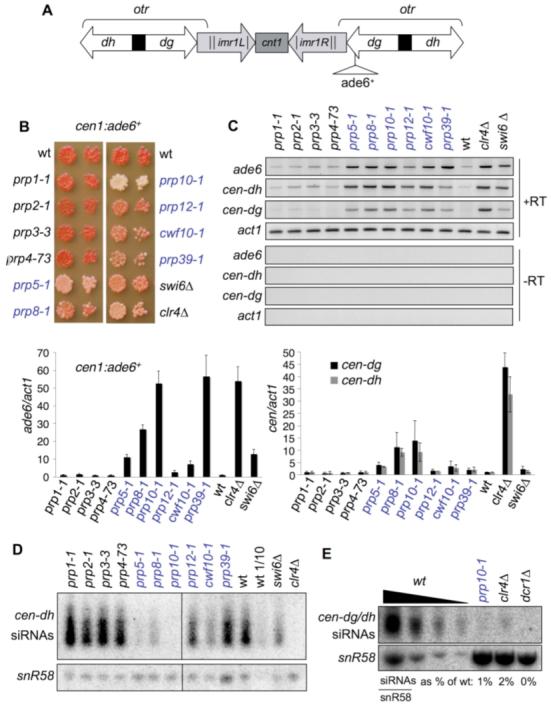
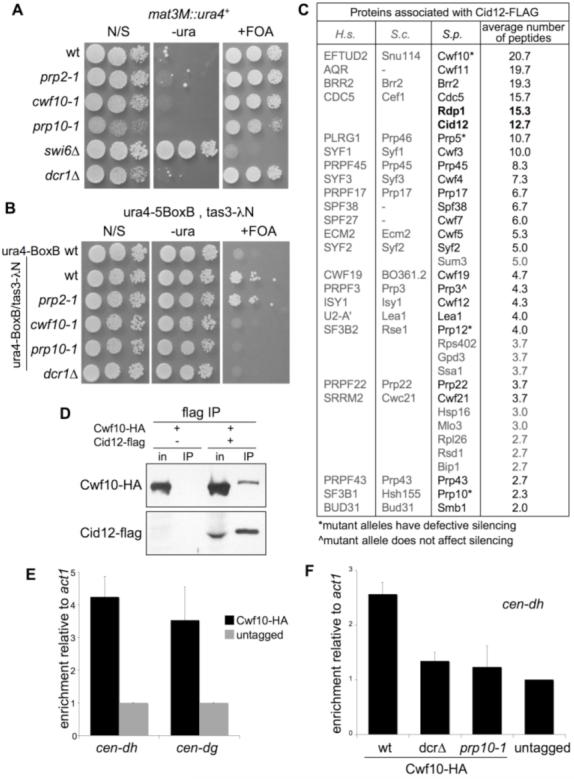
Heterochromatin formation at fission yeast centromeres is directed by RNA interference (RNAi). Noncoding transcripts derived from centromeric repeats are processed into small interfering RNAs (siRNAs) that direct the RNA-induced transcriptional silencing (RITS) effector complex to engage centromere transcripts, resulting in recruitment of the histone H3 lysine 9 methyltransferase Clr4, and hence silencing. We have found that defects in specific splicing factors, but not splicing itself, affect the generation of centromeric siRNAs and consequently centromeric heterochromatin integrity. Moreover, splicing factors physically associate with Cid12, a component of the RNAi machinery, and with centromeric chromatin, consistent with a direct role in RNAi. We propose that spliceosomal complexes provide a platform for siRNA generation and hence facilitate effective centromere repeat silencing.
Figures


References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
