

ZHP-3 acts at crossovers to couple meiotic recombination with synaptonemal complex disassembly and bivalent formation in C. elegans
- PMID: 18949042
- PMCID: PMC2567099
- DOI: 10.1371/journal.pgen.1000235
ZHP-3 acts at crossovers to couple meiotic recombination with synaptonemal complex disassembly and bivalent formation in C. elegans
Erratum in
- PLoS Genet. 2008 Nov;4(11).doi.org/10.1371/annotation/ffbb52bd-7ceb-404c-8c96-93577bf83932
Abstract
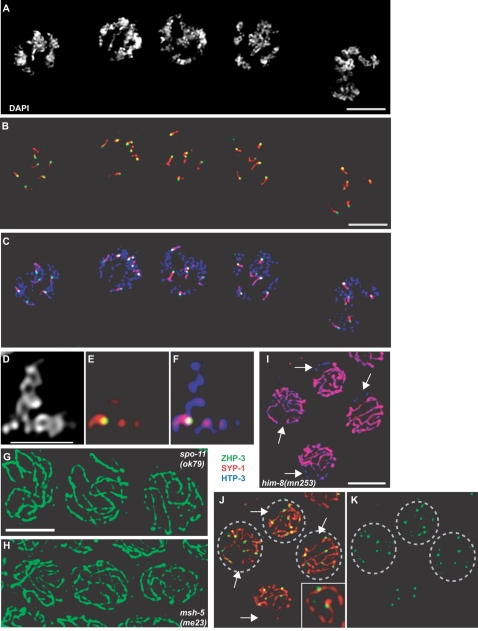
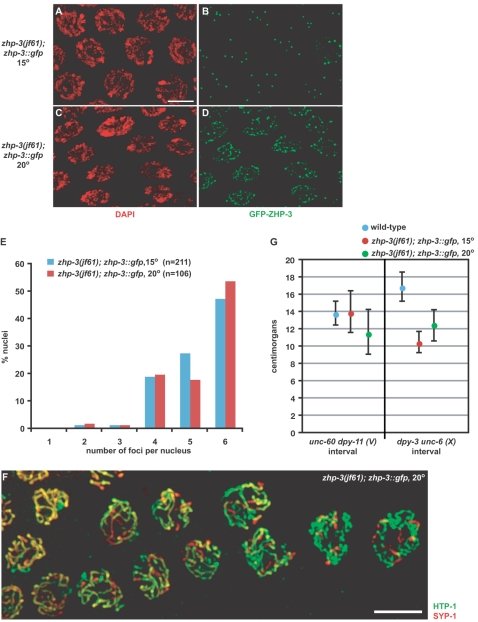
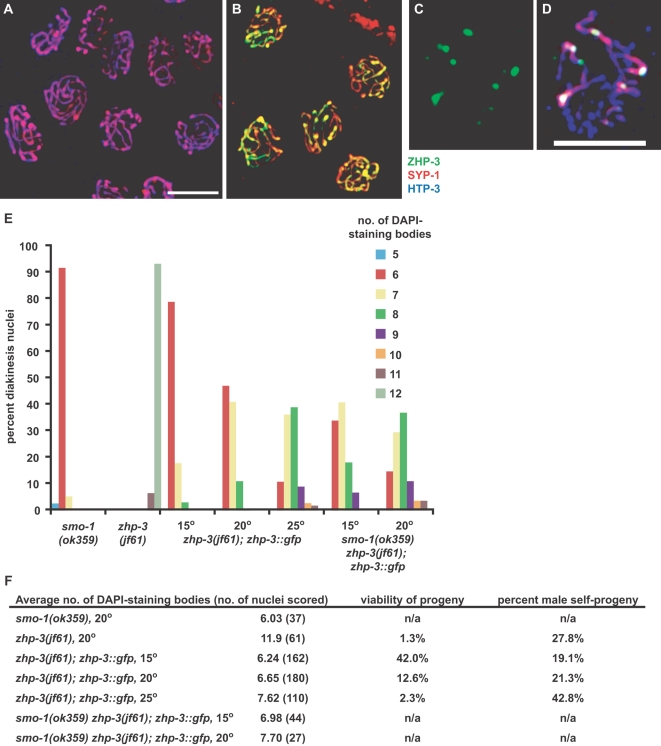
Crossover recombination and the formation of chiasmata normally ensure the proper segregation of homologous chromosomes during the first meiotic division. zhp-3, the Caenorhabditis elegans ortholog of the budding yeast ZIP3 gene, is required for crossover recombination. We show that ZHP-3 protein localization is highly dynamic. At a key transition point in meiotic prophase, the protein shifts from along the length of the synaptonemal complex (SC) to an asymmetric localization on the SC and eventually becomes restricted to foci that mark crossover recombination events. A zhp-3::gfp transgene partially complements a null mutation and reveals a separation of function; although the fusion protein can promote nearly wild-type levels of recombination, aneuploidy among the progeny is high, indicating defects in meiotic chromosome segregation. The structure of bivalents is perturbed in this mutant, suggesting that the chromosome segregation defect results from an inability to properly remodel chromosomes in response to crossovers. smo-1 mutants exhibit phenotypes similar to zhp-3::gfp mutants at higher temperatures, and smo-1; zhp-3::gfp double mutants exhibit more severe meiotic defects than either single mutant, consistent with a role for SUMO in the process of SC disassembly and bivalent differentiation. We propose that coordination of crossover recombination with SC disassembly and bivalent formation reflects a conserved role of Zip3/ZHP-3 in coupling recombination with SC morphogenesis.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures



References
-
- Agarwal S, Roeder GS. Zip3 provides a link between recombination enzymes and synaptonemal complex proteins. Cell. 2000;102:245–255. - PubMed
-
- Borner GV, Kleckner N, Hunter N. Crossover/noncrossover differentiation, synaptonemal complex formation, and regulatory surveillance at the leptotene/zygotene transition of meiosis. Cell. 2004;117:29–45. - PubMed
-
- Fung JC, Rockmill B, Odell M, Roeder GS. Imposition of crossover interference through the nonrandom distribution of synapsis initiation complexes. Cell. 2004;116:795–802. - PubMed
-
- Gill G. SUMO and ubiquitin in the nucleus: different functions, similar mechanisms? Genes Dev. 2004;18:2046–2059. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
