

Review
doi: 10.1080/03602530802431439.
The aldo-keto reductase superfamily and its role in drug metabolism and detoxification
Affiliations
- PMID: 18949601
- PMCID: PMC2663408
- DOI: 10.1080/03602530802431439
Item in Clipboard
Review
The aldo-keto reductase superfamily and its role in drug metabolism and detoxification
Drug Metab Rev.
2008.
Abstract
The aldo-keto reductase (AKR) superfamily comprises enzymes that catalyze redox transformations involved in biosynthesis, intermediary metabolism, and detoxification. Substrates of AKRs include glucose, steroids, glycosylation end-products, lipid peroxidation products, and environmental pollutants. These proteins adopt a (beta/alpha)(8) barrel structural motif interrupted by a number of extraneous loops and helixes that vary between proteins and bring structural identity to individual families. The human AKR family differs from the rodent families. Due to their broad substrate specificity, AKRs play an important role in the phase II detoxification of a large number of pharmaceuticals, drugs, and xenobiotics.
Figures
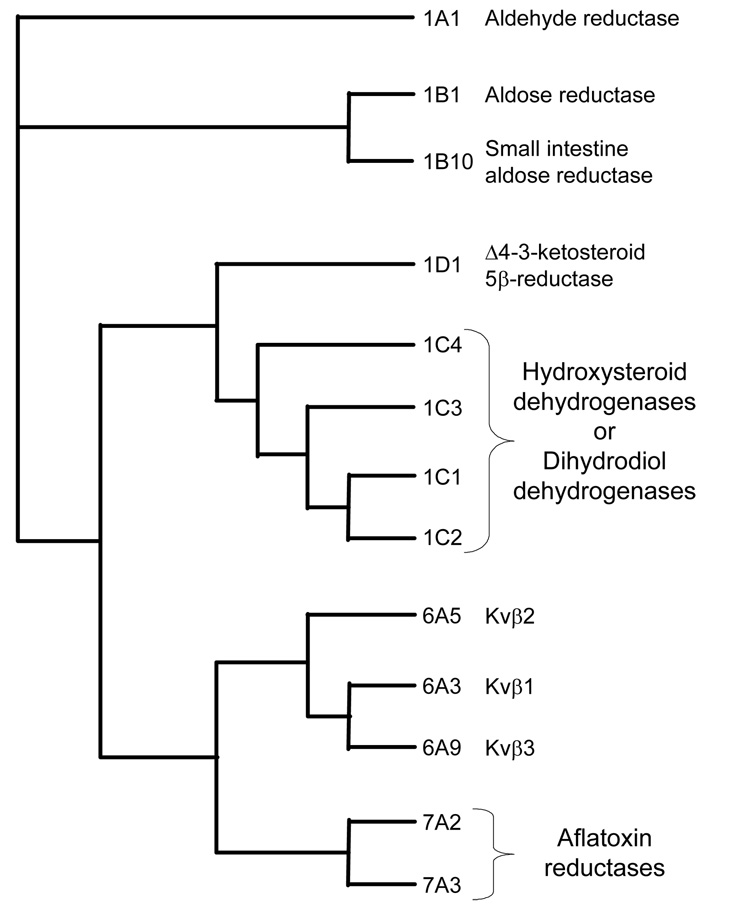
Thirteen well-known and characterized human proteins belong to 3 AKR families.
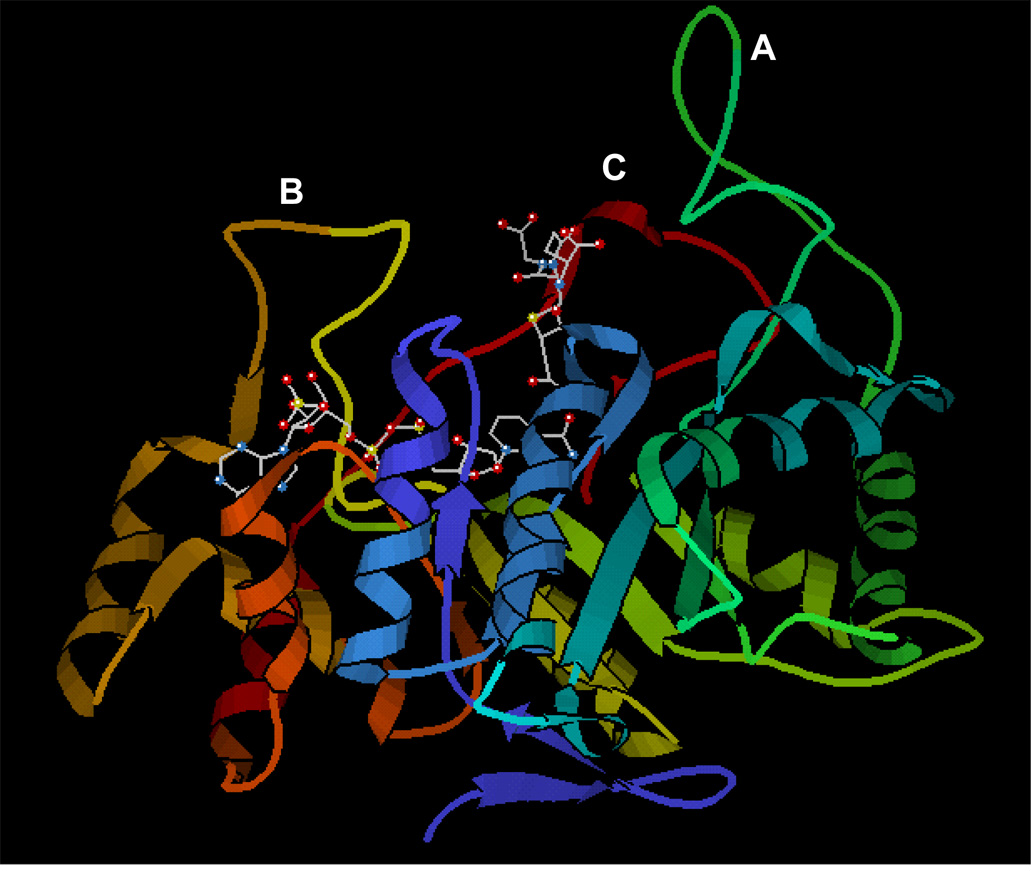
The structure was downloaded from RCSB Protein Data Bank (ID # 2f2k). The ribbon drawing is a side-view representation of the protein with NADPH bound to the active site. The active-site loops are marked - A, B and C. The ball and stick structure at loop B is the NADPH molecule and the structure at loop C is the glutathione analog 1,2-dicarboxyethyl glutathione, which binds to the substrate binding site of the enzyme.
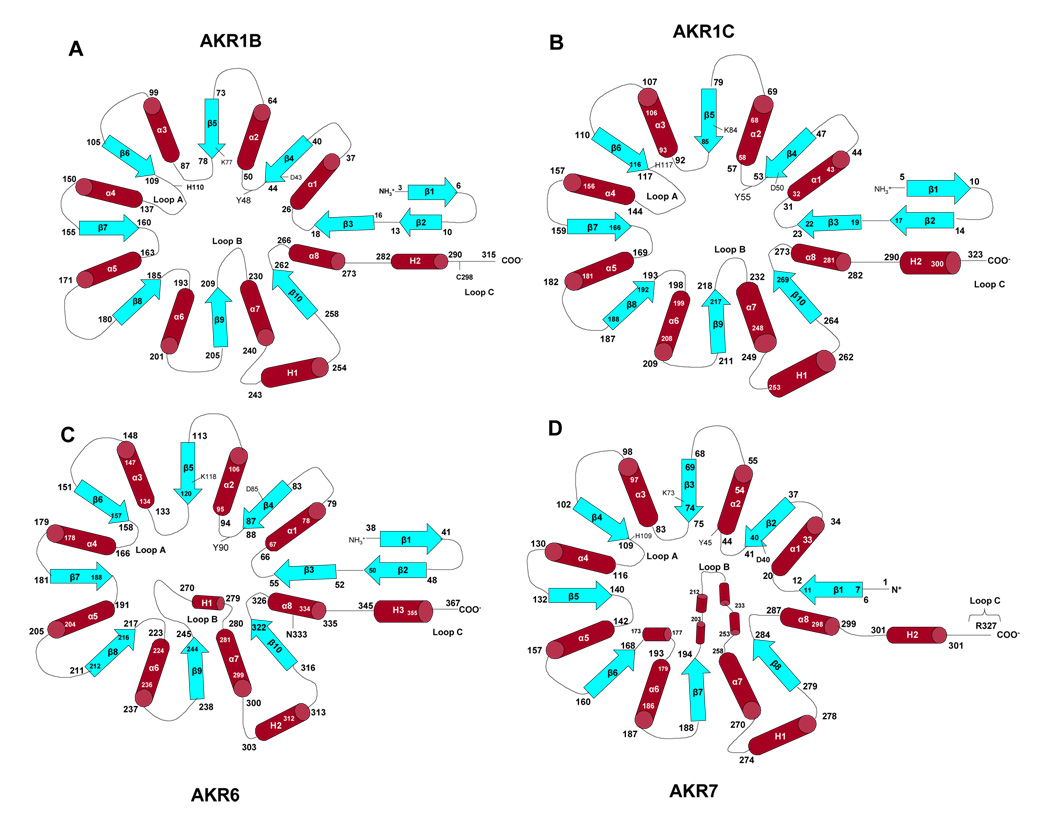
Topological representations of the amino acid sequence of AKR1B (A), AKR1C (B), AKR6 (C), and AKR7 (D). The β-strands (blue) are represented as arrows and the α-helices (red) as cylinders. Each major strand and loop is number. Connecting random coils are shown as extended loops. The position of the active site residues in sequence is indicated. Note the shorter loop A and the extra helices in AKR6 and 7 (C and D).
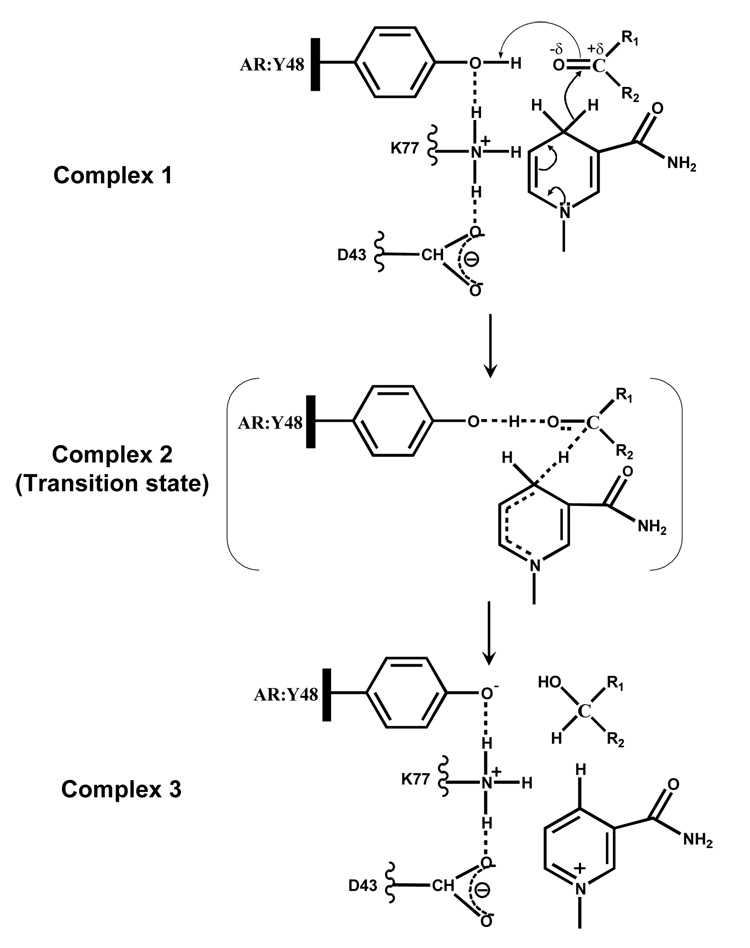
In complex 1, active site Tyr-48 (AKR1B1 numbering) is shown to form a hydrogen bond with the substrate carbonyl resulting in the carbonyl polarization and accelerating the hydride transfer of the pro-R hydrogen from the nicotinamide ring of NADPH to the carbonyl carbon of the substrate. The hydrogen bond network provided by Lys-77 and the Asp-43 serves to lower the pKa of tyrosine making the proton transfer easier. Complex 2 shows a transition state in which the polarization at the carbonyl is quenched by the proton transferred from the protein tyrosine and a concerted hydride transfer to the carbonyl carbon. The reduced carbonyl then dissociates from the acid-base catalyst and a net charge on the tyrosinate anion is stabilized by the hydrogen-bonding network (complex 3).
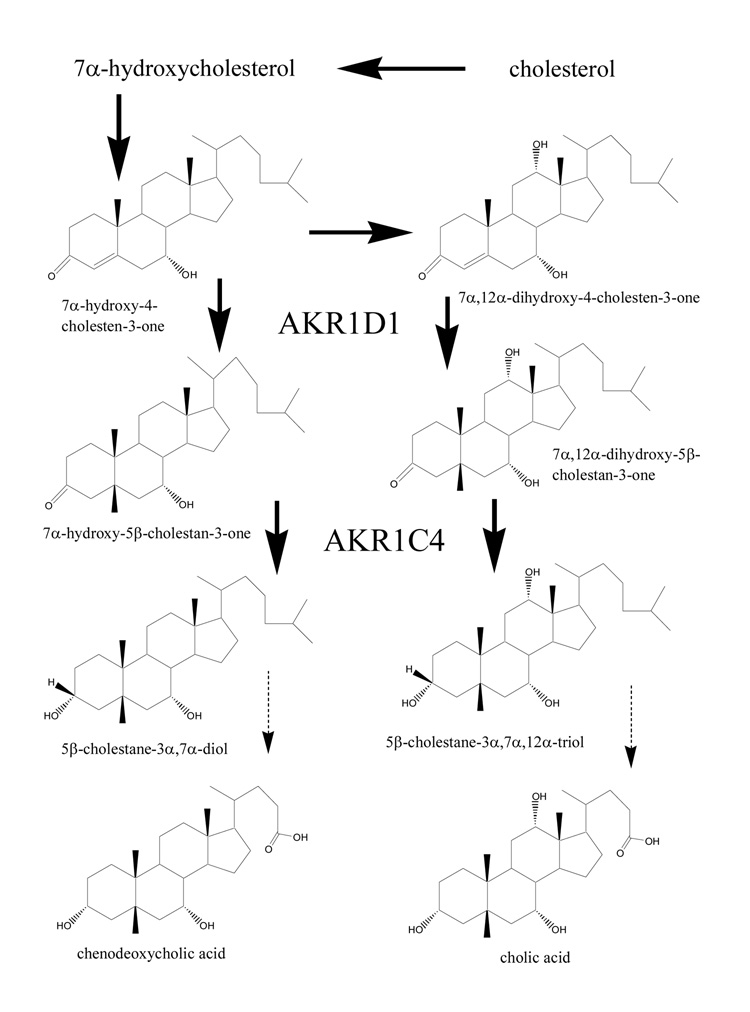
The AKR 1D1 and 1C4 catalyze two consecutive steps in the synthesis of cholic and deoxycholic acids.
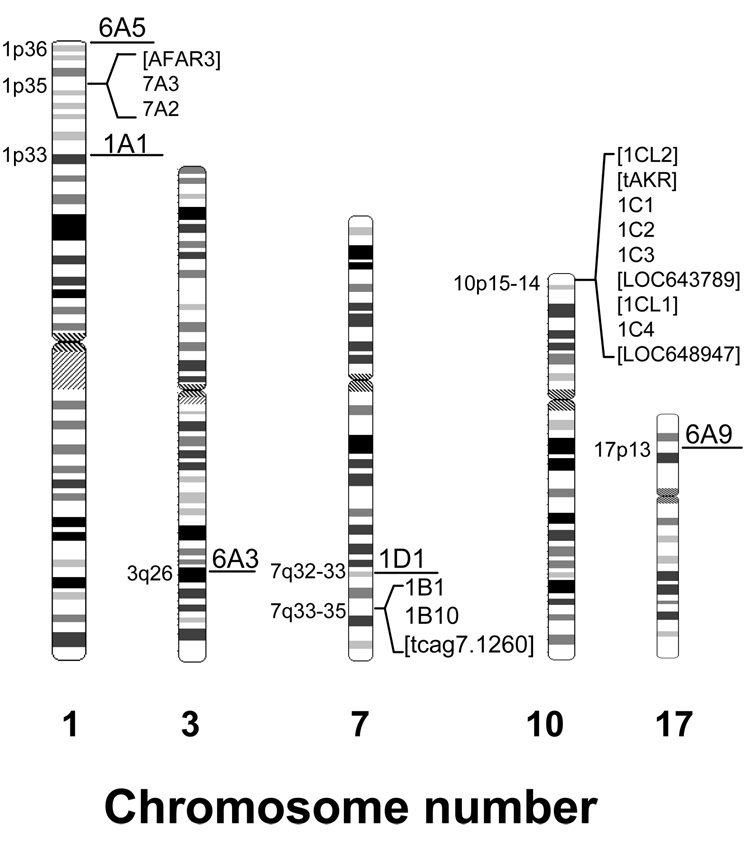
With the exception of AKR6, the AKR families form clusters on the same chromosome. Genes for which protein product has not been demonstrated are shown in square brackets.
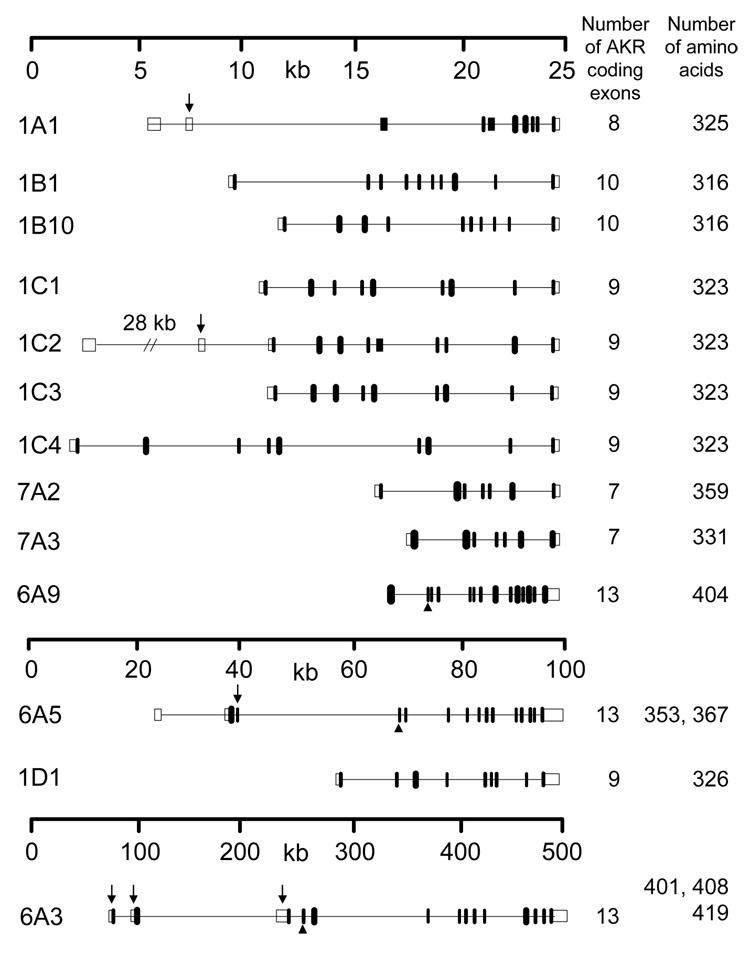
Open boxes represent the noncoding exons and filled boxes indicate the coding exons. Arrows point to the exons subject to alternative splicing. The arrowheads show the start of the AKR domain in the Kvβ (AKR6) family. The number of exons coding for the AKR domain and the amino acid length of the corresponding protein are shown in adjacent columns.
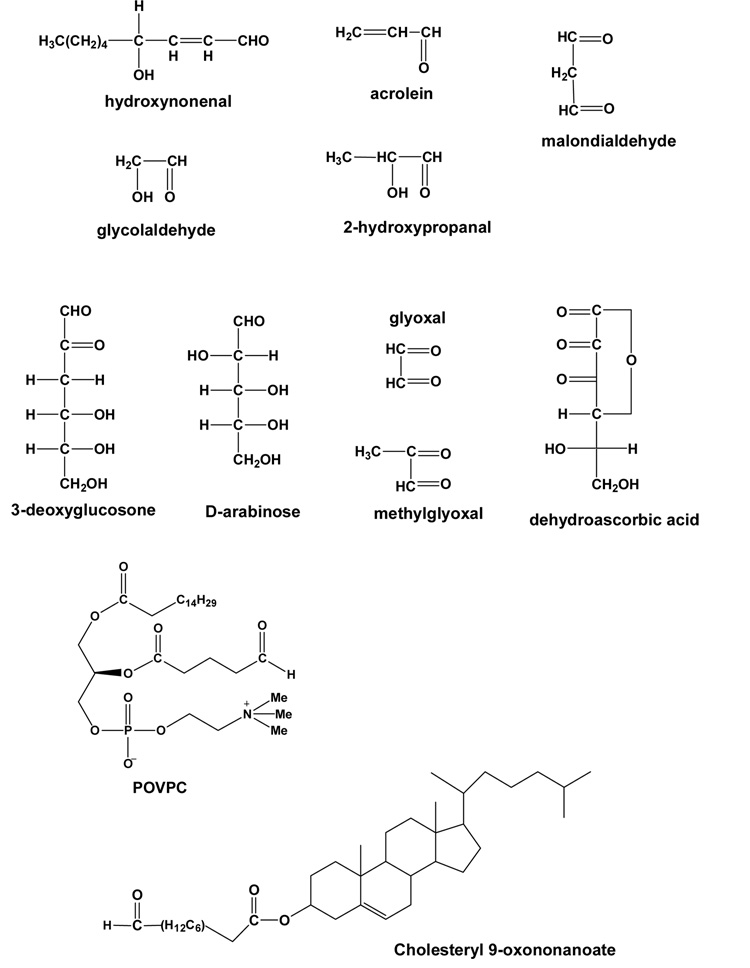
Chemical structures of the endogenous carbonyls identified as lipid peroxidation products, or carbohydrate-derived advanced glycosylation end-product (AGE) precursors are shown. At the bottom are representative structures of core aldehydes generated during the oxidation of phospholipids (e.g. 1-palmitoyl-2-arachidonyl phosphatidylcholine) or cholesterol esters containing unsaturated fatty acid side chains. POVPC: 1-palmitoyl-2-oxovaleroyl phosphatidyl choline.
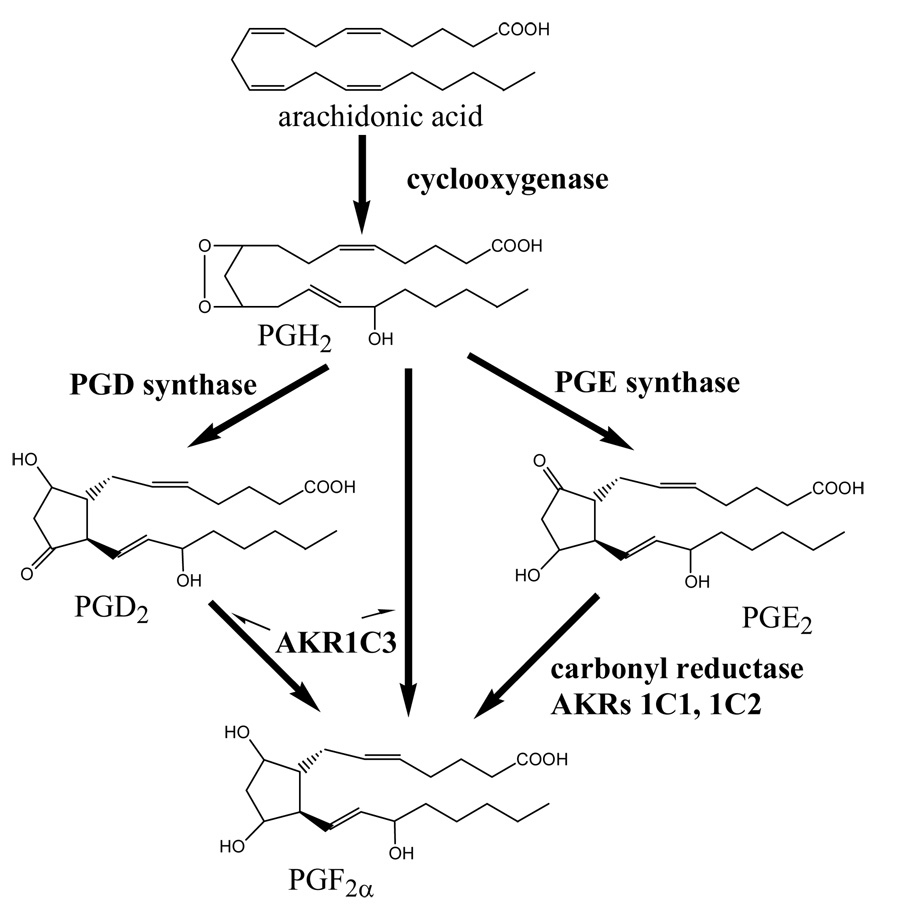
Members of the AKR family catalyze the transformation of PGH2, PGD2 and PGE2 into PGF2.
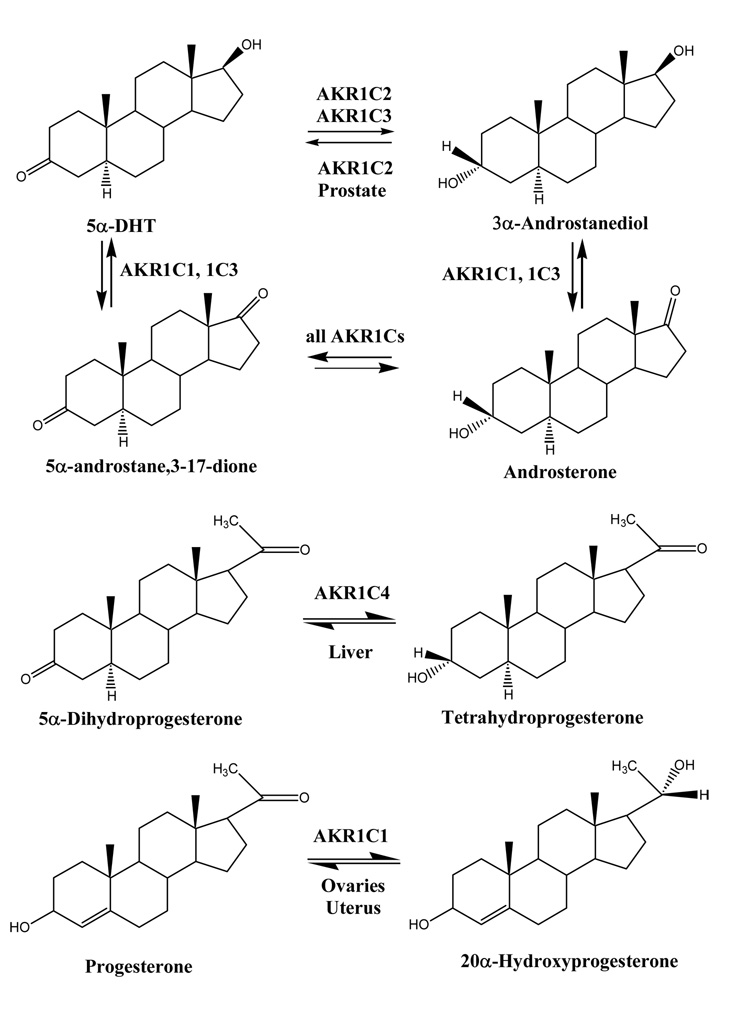
Several AKR1C enzymes catalyze oxidation-reduction reactions at the 3, 17, and 20 positions of the steroid molecule.
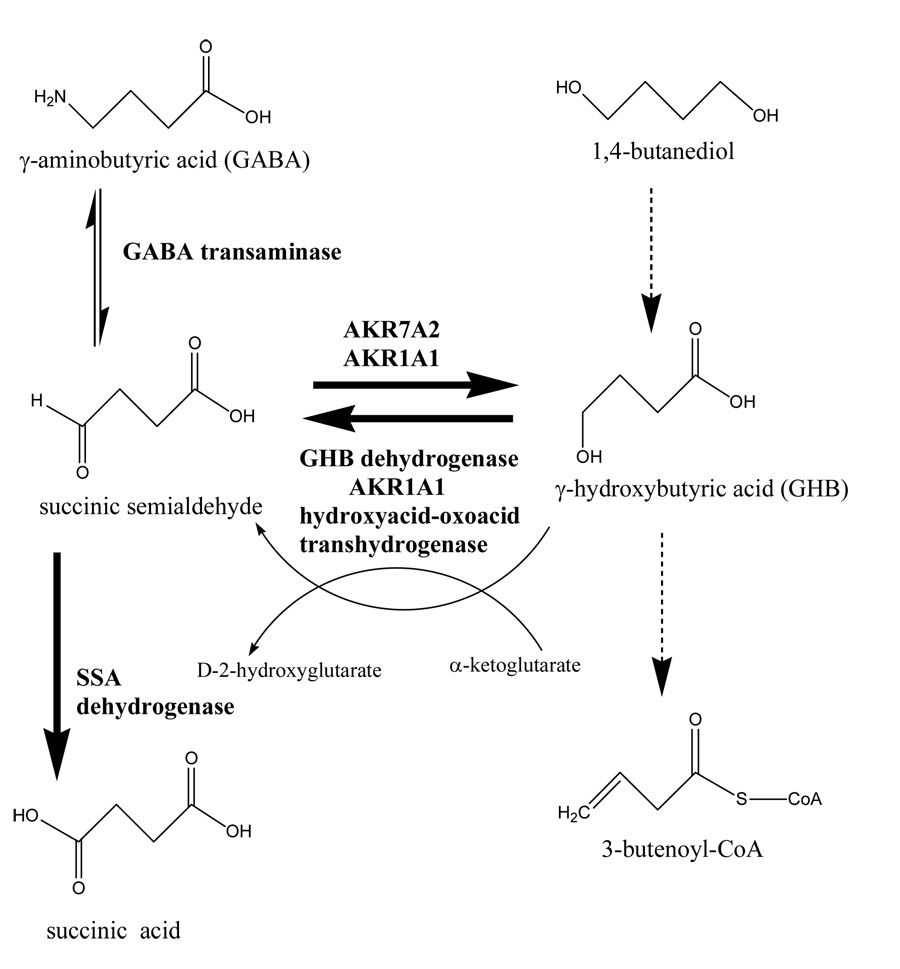
Members of AKR families 1 and 7 catalyze the interconversion between succinic semialdehyde and γ-hydroxybutyrate (GHB).
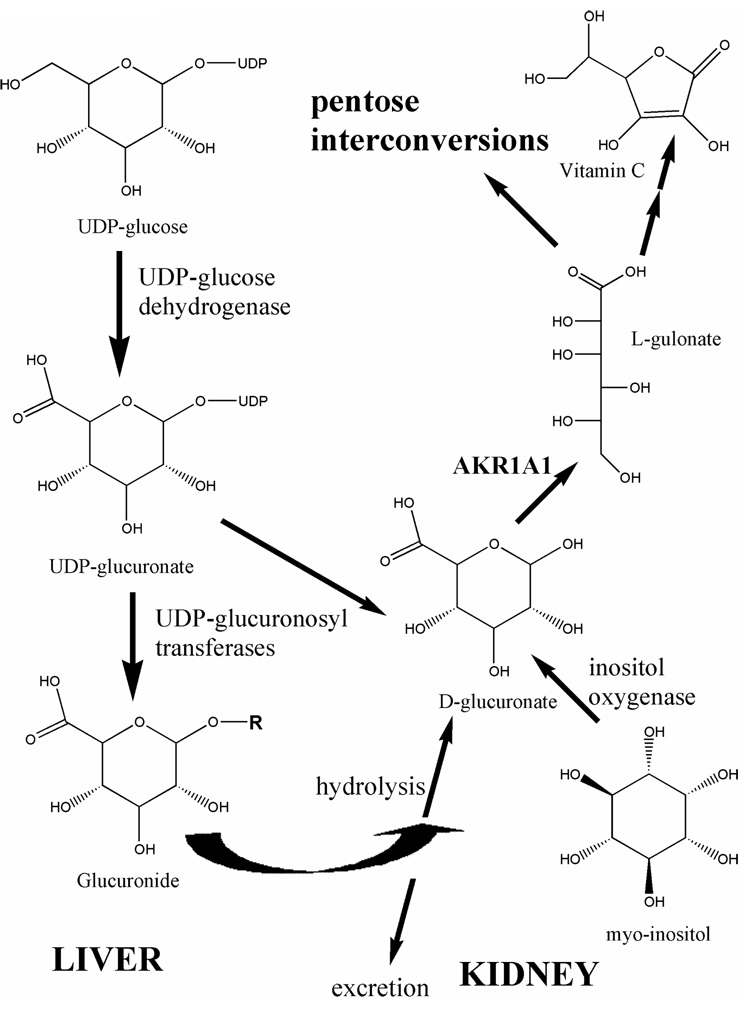
AKR1A catalyzes the reduction of D-glucuronate to L-gulonate.
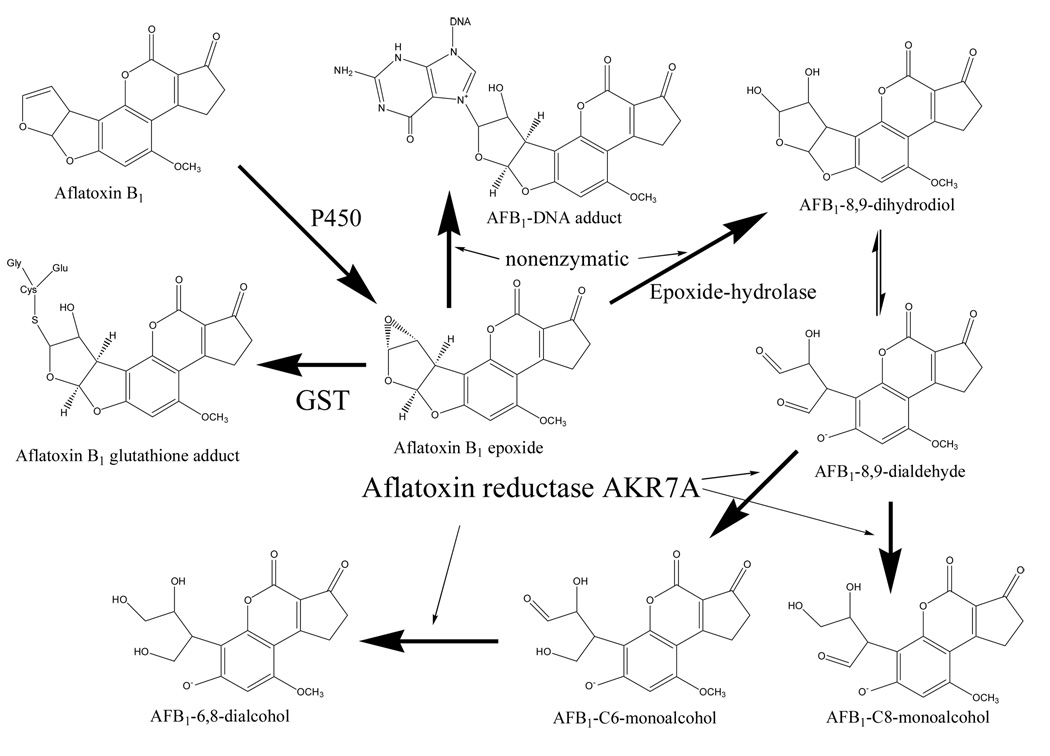
Aflatoxin reductases of the AKR7 family catalyze the reduction of AFB1 dialdehyde, the cytotoxic intermediate of aflatoxin B1 activation, to mono-and dialcohols as indicated.
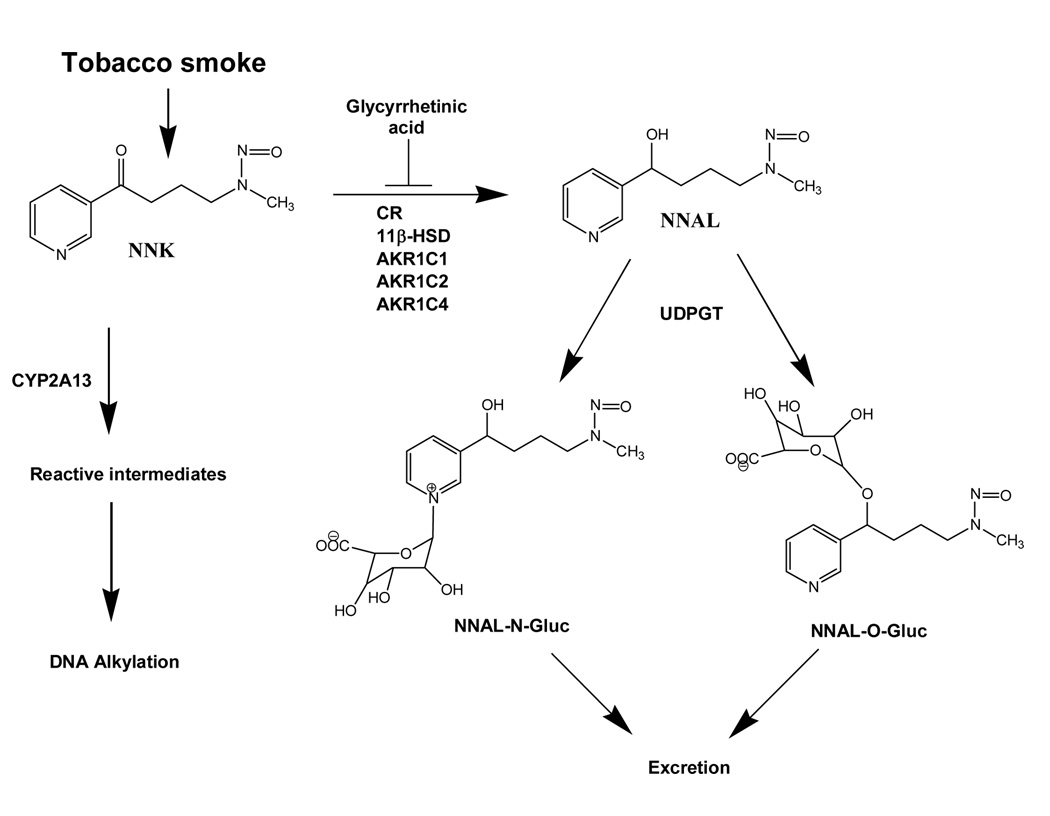
NNK can be oxygenated by cytochromes P450 or reduced by several enzymes of the AKR and HSD families to form nitrosamine alcohol (NNAL). Reduction represents detoxification pathway as NNAL is glucuronydated by uridine diphosphate glucuronosyl transferases (UDPGT) and is subsequently excreted.
References
-
- Aimond F, Kwak SP, Rhodes KJ, Nerbonne JM. Accessory kv beta 1 subunits differentially modulate the functional expression of voltage-gated K+ channels in mouse ventricular myocytes. Circulation Research. 2005;96:451–458. - PubMed
-
- Atalla A, Breyer-Pfaff U, Maser E. Purification and characterization of oxidoreductases-catalyzing carbonyl reduction of the tobacco-specific nitrosamine 4-methylnitrosamino-1-(3-pyridyl)-1-butanone (NNK) in human liver cytosol. Xenobiotica. 2000;30:755–769. - PubMed
-
- Atalla A, Maser E. Characterization of enzymes participating in carbonyl reduction of 4-methylnitrosamino-1-(3-pyridyl)-1-butanone (NNK) in human placenta. Chemico-Biological Interactions. 2001;130:737–748. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical