

De novo DNA methylation promoted by G9a prevents reprogramming of embryonically silenced genes
- PMID: 18953337
- PMCID: PMC2581722
- DOI: 10.1038/nsmb.1476
De novo DNA methylation promoted by G9a prevents reprogramming of embryonically silenced genes
Abstract
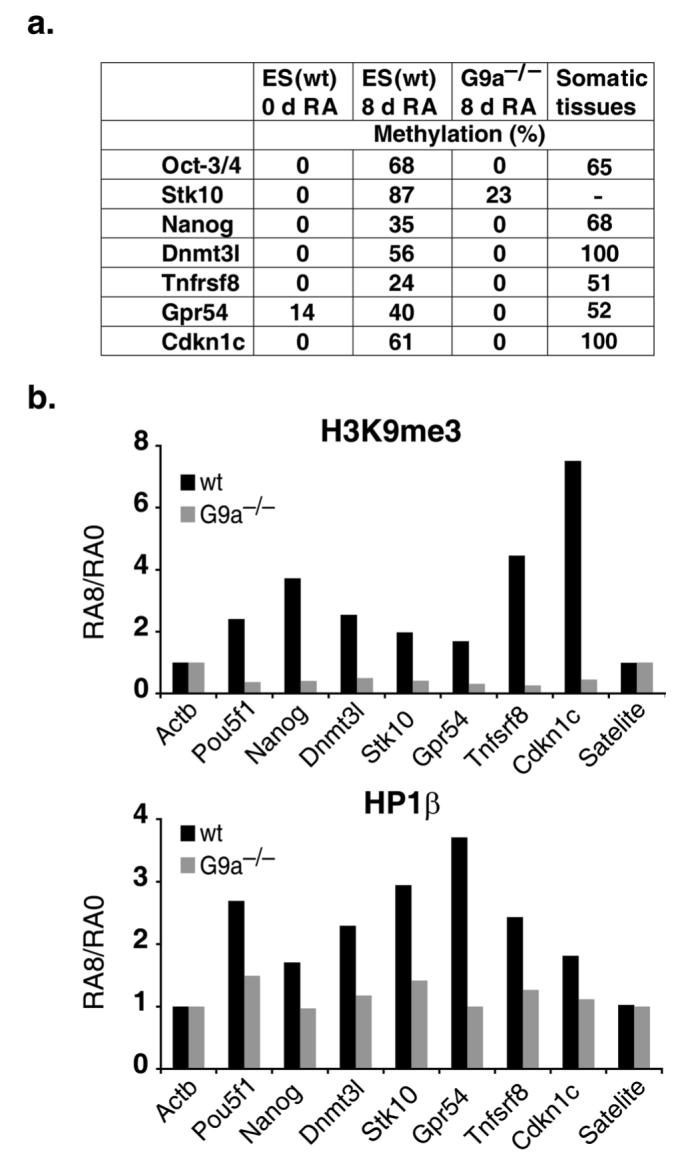
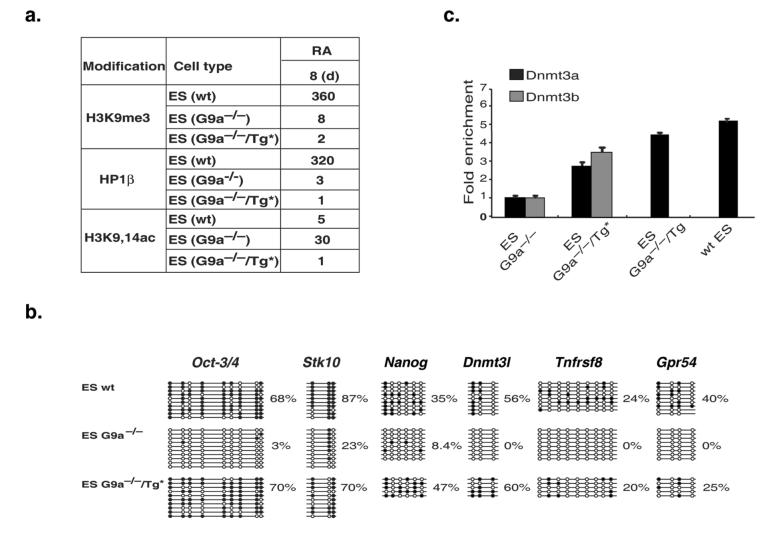
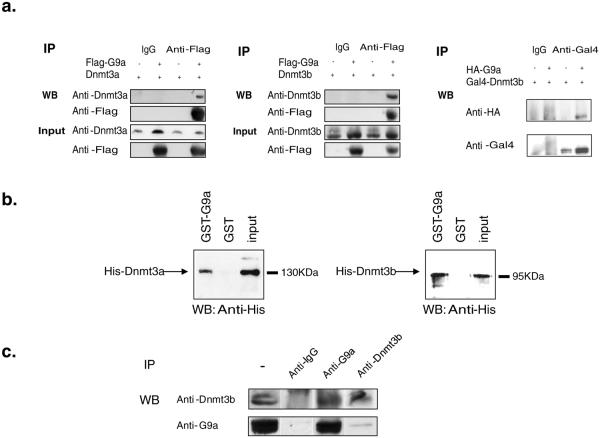
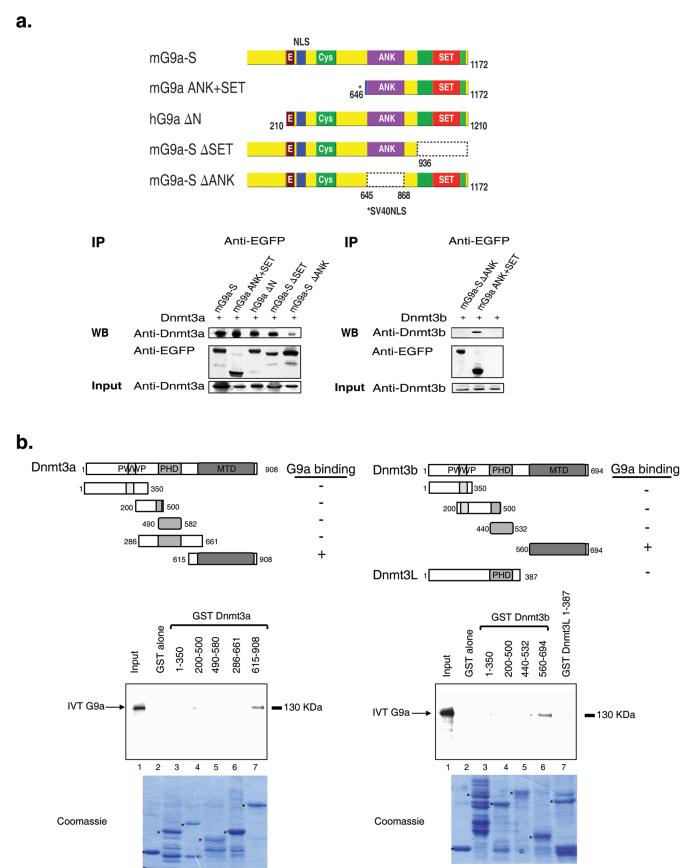
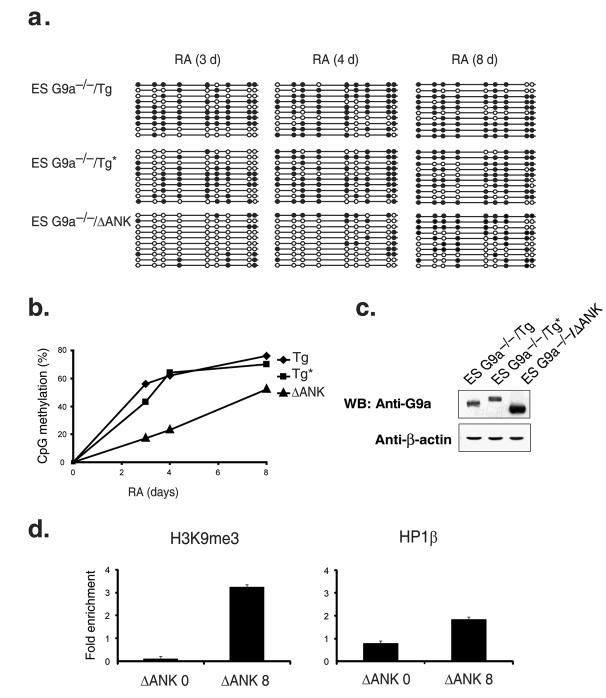
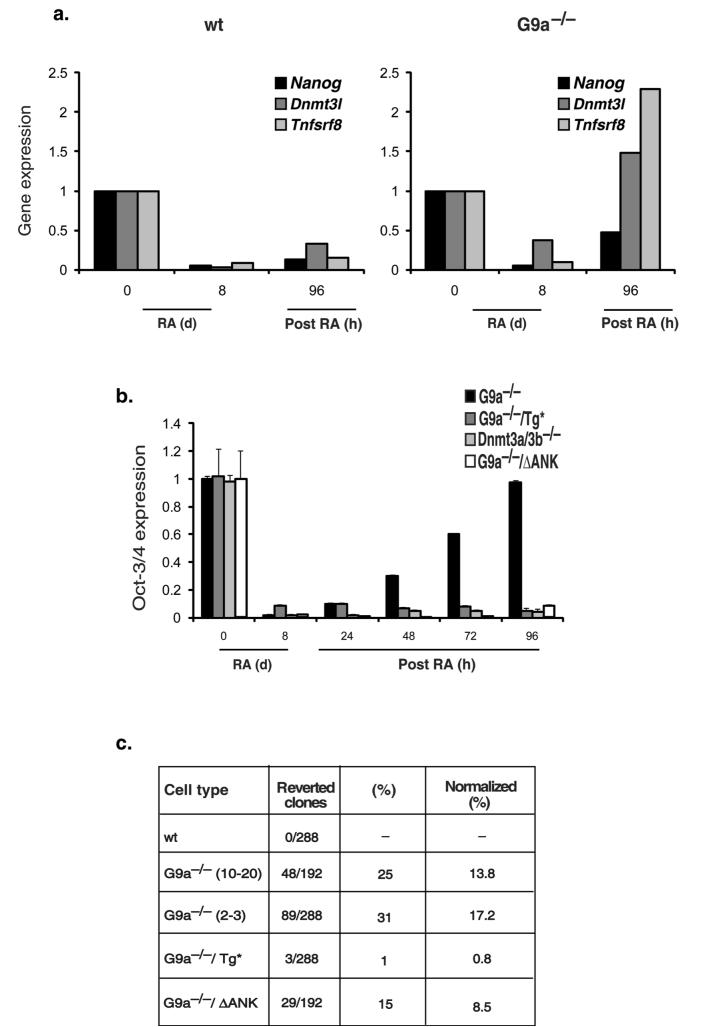
The pluripotency-determining gene Oct3/4 (also called Pou5f1) undergoes postimplantation silencing in a process mediated by the histone methyltransferase G9a. Microarray analysis now shows that this enzyme may operate as a master regulator that inactivates numerous early-embryonic genes by bringing about heterochromatinization of methylated histone H3K9 and de novo DNA methylation. Genetic studies in differentiating embryonic stem cells demonstrate that a point mutation in the G9a SET domain prevents heterochromatinization but still allows de novo methylation, whereas biochemical and functional studies indicate that G9a itself is capable of bringing about de novo methylation through its ankyrin domain, by recruiting Dnmt3a and Dnmt3b independently of its histone methyltransferase activity. These modifications seem to be programmed for carrying out two separate biological functions: histone methylation blocks target-gene reactivation in the absence of transcriptional repressors, whereas DNA methylation prevents reprogramming to the undifferentiated state.
Figures
References
-
- Surani MA, Hayashi K, Hajkova P. Genetic and epigenetic regulators of pluripotency. Cell. 2007;128:747–762. - PubMed
-
- Reik W. Stability and flexibility of epigenetic gene regulation in mammalian development. Nature. 2007;447:425–432. - PubMed
-
- Niwa H. Open conformation chromatin and pluripotency. Genes Dev. 2007;21:2671–2676. - PubMed
-
- Niwa H, Miyazaki J, Smith AG. Quantitative expression of Oct-3/4 defines differentiation, dedifferentiation or self-renewal of ES cells. Nature Genet. 2000;24:372–376. - PubMed
-
- Chambers I, et al. Functional expression cloning of nanog, a pluripotency sustaining factor in embryonic stem cells. Cell. 2003;113:643–655. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
