

Telomeric expression sites are highly conserved in Trypanosoma brucei
- PMID: 18953401
- PMCID: PMC2567434
- DOI: 10.1371/journal.pone.0003527
Telomeric expression sites are highly conserved in Trypanosoma brucei
Abstract
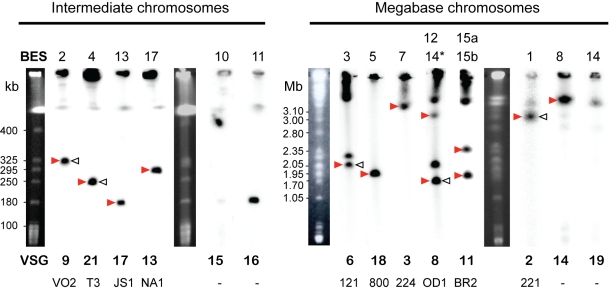
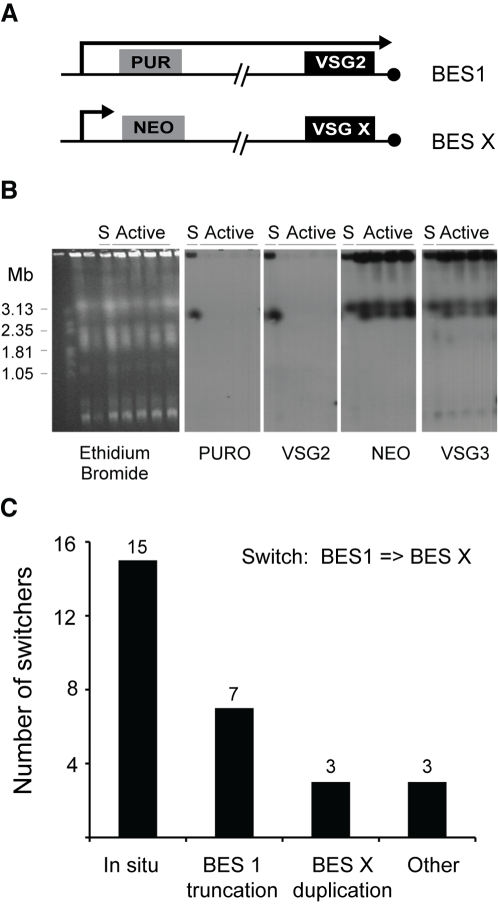
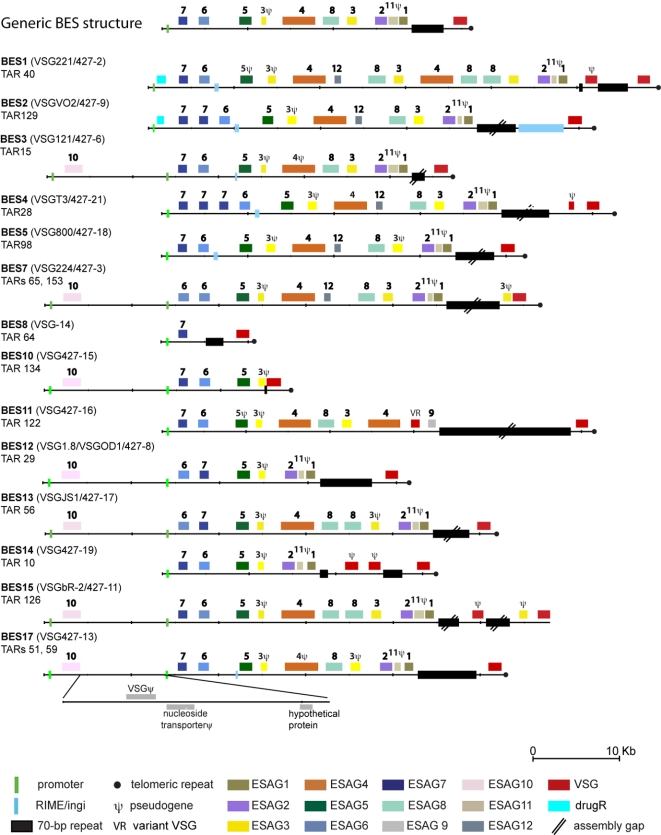
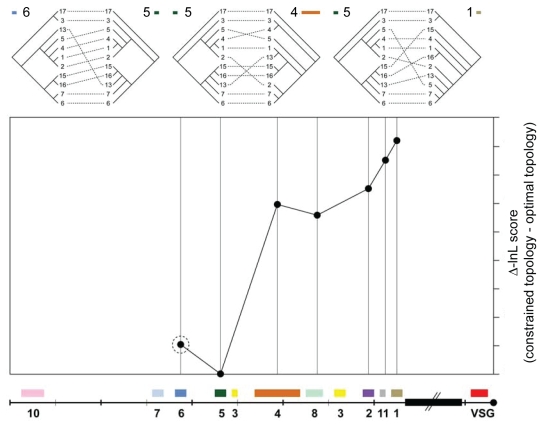
Subtelomeric regions are often under-represented in genome sequences of eukaryotes. One of the best known examples of the use of telomere proximity for adaptive purposes are the bloodstream expression sites (BESs) of the African trypanosome Trypanosoma brucei. To enhance our understanding of BES structure and function in host adaptation and immune evasion, the BES repertoire from the Lister 427 strain of T. brucei were independently tagged and sequenced. BESs are polymorphic in size and structure but reveal a surprisingly conserved architecture in the context of extensive recombination. Very small BESs do exist and many functioning BESs do not contain the full complement of expression site associated genes (ESAGs). The consequences of duplicated or missing ESAGs, including ESAG9, a newly named ESAG12, and additional variant surface glycoprotein genes (VSGs) were evaluated by functional assays after BESs were tagged with a drug-resistance gene. Phylogenetic analysis of constituent ESAG families suggests that BESs are sequence mosaics and that extensive recombination has shaped the evolution of the BES repertoire. This work opens important perspectives in understanding the molecular mechanisms of antigenic variation, a widely used strategy for immune evasion in pathogens, and telomere biology.
Conflict of interest statement
Figures
References
-
- Mefford HC, Trask BJ. The complex structure and dynamic evolution of human subtelomeres. Nat Rev Genet. 2002;3:91–102. - PubMed
-
- Horn D, Barry JD. The central roles of telomeres and subtelomeres in antigenic variation in African trypanosomes. Chromosome Res. 2005;13:525–533. - PubMed
-
- Pryde FE, Gorham HC, Louis EJ. Chromosome ends: all the same under their caps. Curr Opin Genet Dev. 1997;7:822–828. - PubMed
-
- Pays E. Regulation of antigen gene expression in Trypanosoma brucei. Trends Parasitol. 2005;21:517–520. - PubMed
