

Memo-RhoA-mDia1 signaling controls microtubules, the actin network, and adhesion site formation in migrating cells
- PMID: 18955552
- PMCID: PMC2575782
- DOI: 10.1083/jcb.200805107
Memo-RhoA-mDia1 signaling controls microtubules, the actin network, and adhesion site formation in migrating cells
Abstract
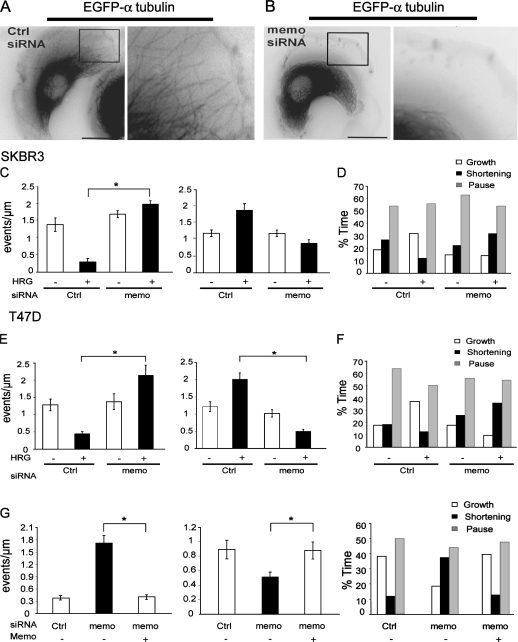
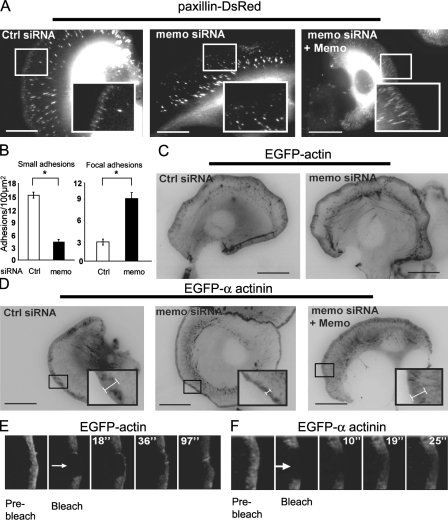
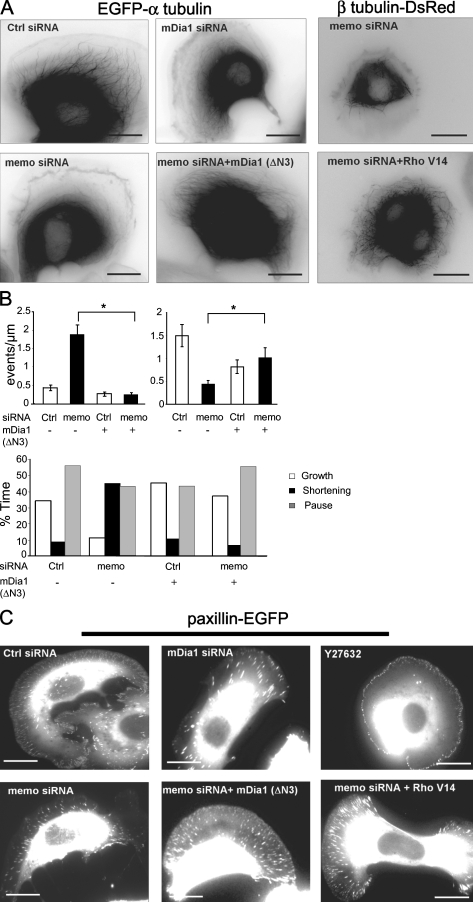
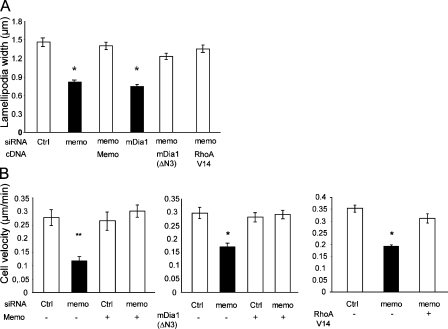
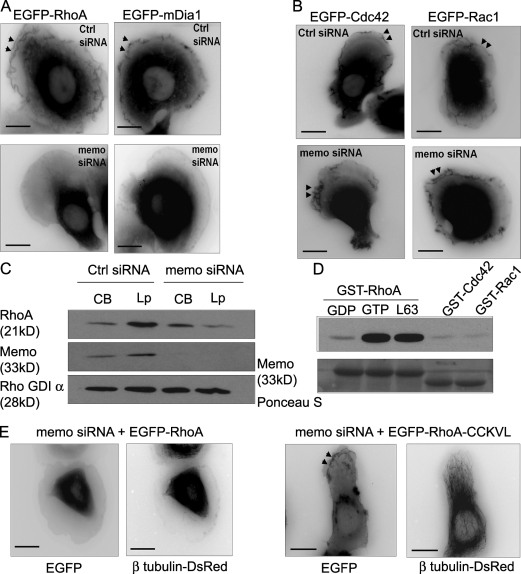
Actin assembly at the cell front drives membrane protrusion and initiates the cell migration cycle. Microtubules (MTs) extend within forward protrusions to sustain cell polarity and promote adhesion site turnover. Memo is an effector of the ErbB2 receptor tyrosine kinase involved in breast carcinoma cell migration. However, its mechanism of action remained unknown. We report in this study that Memo controls ErbB2-regulated MT dynamics by altering the transition frequency between MT growth and shortening phases. Moreover, although Memo-depleted cells can assemble the Rac1-dependent actin meshwork and form lamellipodia, they show defective localization of lamellipodial markers such as alpha-actinin-1 and a reduced number of short-lived adhesion sites underlying the advancing edge of migrating cells. Finally, we demonstrate that Memo is required for the localization of the RhoA guanosine triphosphatase and its effector mDia1 to the plasma membrane and that Memo-RhoA-mDia1 signaling coordinates the organization of the lamellipodial actin network, adhesion site formation, and MT outgrowth within the cell leading edge to sustain cell motility.
Figures
References
-
- Bear, J.E., T.M. Svitkina, M. Krause, D.A. Schafer, J.J. Loureiro, G.A. Strasser, I.V. Maly, O.Y. Chaga, J.A. Cooper, G.G. Borisy, and F.B. Gertler. 2002. Antagonism between Ena/VASP proteins and actin filament capping regulates fibroblast motility. Cell. 109:509–521. - PubMed
-
- Burridge, K., and K. Wennerberg. 2004. Rho and Rac take center stage. Cell. 116:167–179. - PubMed
-
- Ishizaki, T., Y. Morishima, M. Okamoto, T. Furuyashiki, T. Kato, and S. Narumiya. 2001. Coordination of microtubules and the actin cytoskeleton by the Rho effector mDia1. Nat. Cell Biol. 3:8–14. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
