

Segregation and activation of myosin IIB creates a rear in migrating cells
- PMID: 18955554
- PMCID: PMC2575793
- DOI: 10.1083/jcb.200806030
Segregation and activation of myosin IIB creates a rear in migrating cells
Abstract
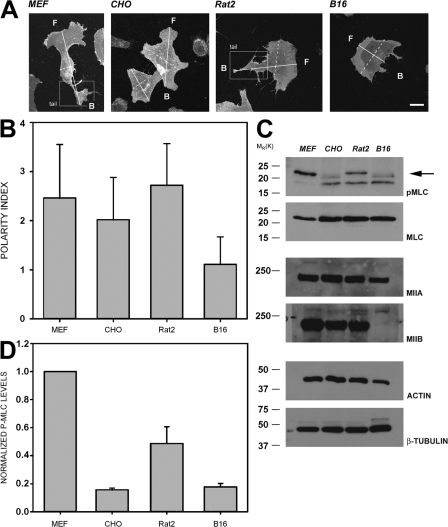
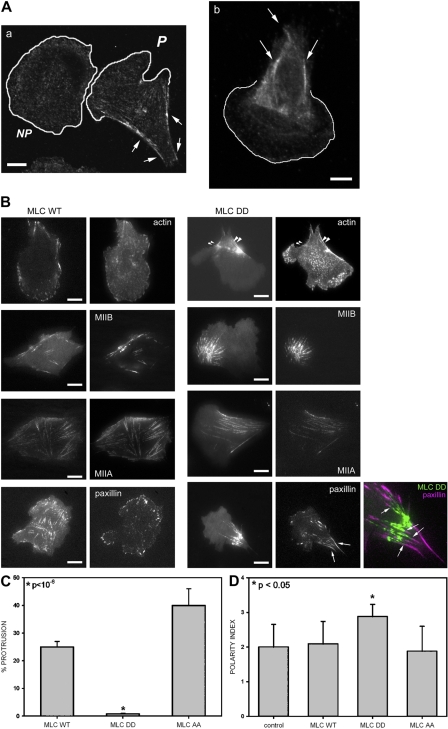
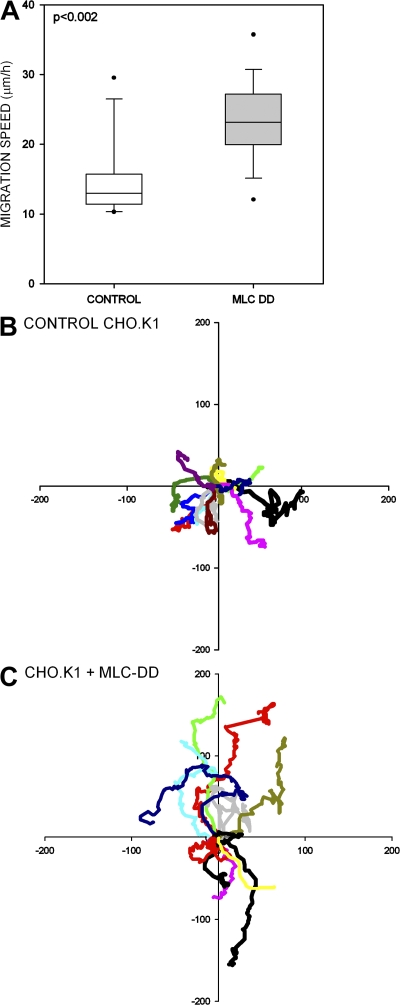
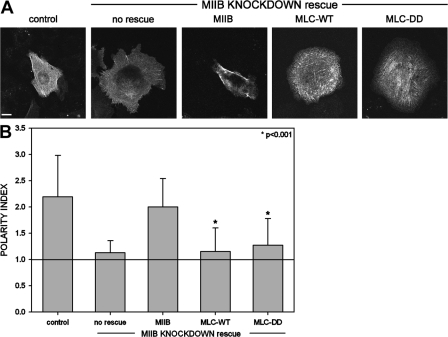
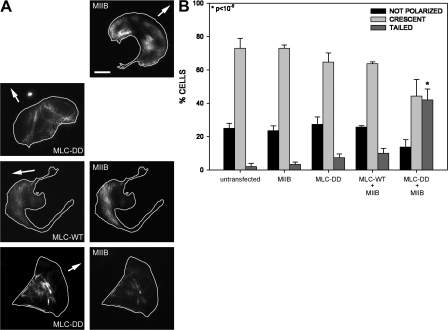
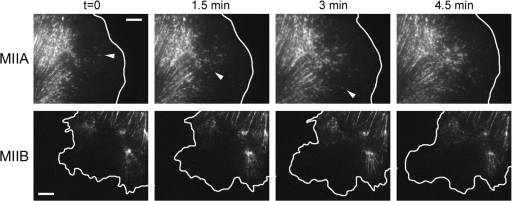
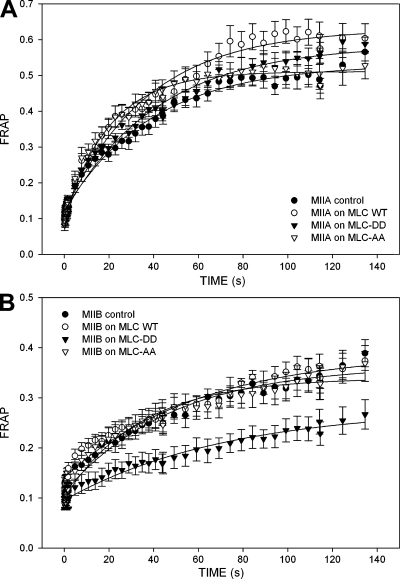
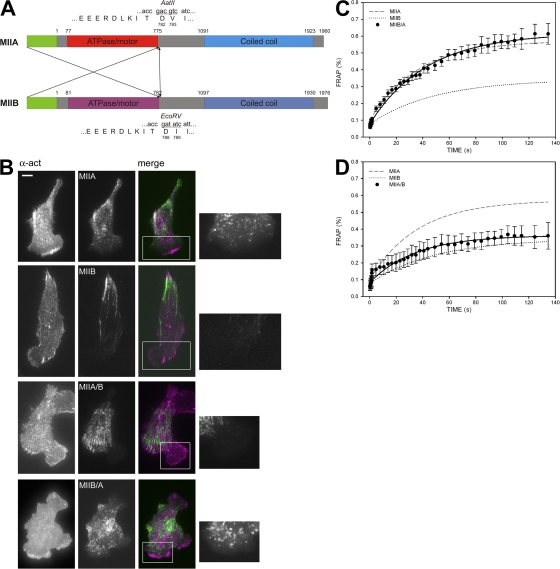
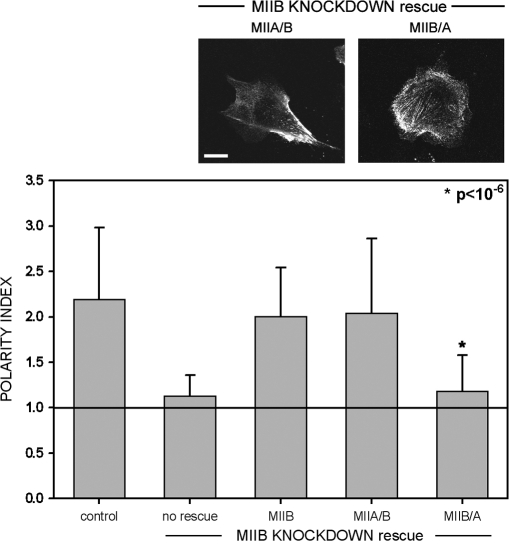
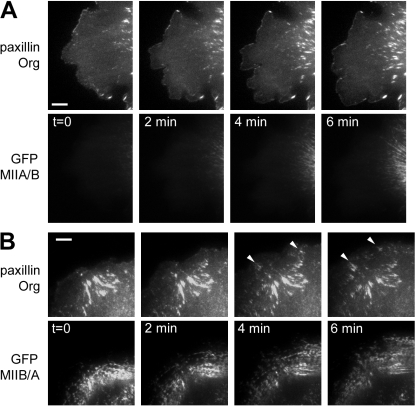
We have found that MLC-dependent activation of myosin IIB in migrating cells is required to form an extended rear, which coincides with increased directional migration. Activated myosin IIB localizes prominently at the cell rear and produces large, stable actin filament bundles and adhesions, which locally inhibit protrusion and define the morphology of the tail. Myosin IIA forms de novo filaments away from the myosin IIB-enriched center and back to form regions that support protrusion. The positioning and dynamics of myosin IIA and IIB depend on the self-assembly regions in their coiled-coil C terminus. COS7 and B16 melanoma cells lack myosin IIA and IIB, respectively; and show isoform-specific front-back polarity in migrating cells. These studies demonstrate the role of MLC activation and myosin isoforms in creating a cell rear, the segregation of isoforms during filament assembly and their differential effects on adhesion and protrusion, and a key role for the noncontractile region of the isoforms in determining their localization and function.
Figures
References
-
- Bao, J., S.S. Jana, and R.S. Adelstein. 2005. Vertebrate nonmuscle myosin II isoforms rescue small interfering RNA-induced defects in COS-7 cell cytokinesis. J. Biol. Chem. 280:19594–19599. - PubMed
-
- Burridge, K., and M. Chrzanowska-Wodnicka. 1996. Focal adhesions, contractility, and signaling. Annu. Rev. Cell Dev. Biol. 12:463–518. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
