

Rapamycin differentially inhibits S6Ks and 4E-BP1 to mediate cell-type-specific repression of mRNA translation
- PMID: 18955708
- PMCID: PMC2582304
- DOI: 10.1073/pnas.0809136105
Rapamycin differentially inhibits S6Ks and 4E-BP1 to mediate cell-type-specific repression of mRNA translation
Abstract
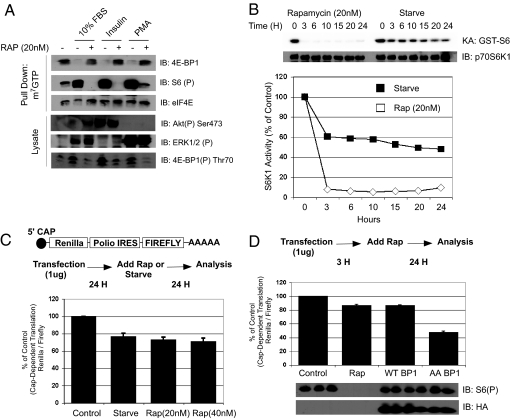
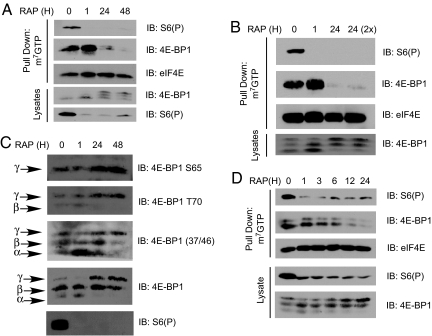
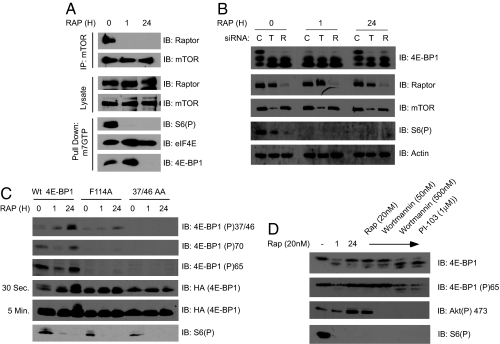
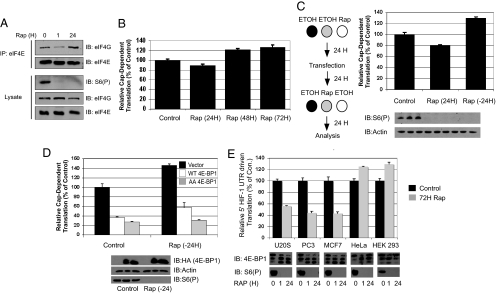
The mammalian translational initiation machinery is a tightly controlled system that is composed of eukaryotic initiation factors, and which controls the recruitment of ribosomes to mediate cap-dependent translation. Accordingly, the mTORC1 complex functionally controls this cap-dependent translation machinery through the phosphorylation of its downstream substrates 4E-BPs and S6Ks. It is generally accepted that rapamycin, a specific inhibitor of mTORC1, is a potent translational repressor. Here we report the unexpected discovery that rapamycin's ability to regulate cap-dependent translation varies significantly among cell types. We show that this effect is mechanistically caused by rapamycin's differential effect on 4E-BP1 versus S6Ks. While rapamycin potently inhibits S6K activity throughout the duration of treatment, 4E-BP1 recovers in phosphorylation within 6 h despite initial inhibition (1-3 h). This reemerged 4E-BP1 phosphorylation is rapamycin-resistant but still requires mTOR, Raptor, and mTORC1's activity. Therefore, these results explain how cap-dependent translation can be maintained in the presence of rapamycin. In addition, we have also defined the condition by which rapamycin can control cap-dependent translation in various cell types. Finally, we show that mTOR catalytic inhibitors are effective inhibitors of the rapamycin-resistant phenotype.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Gingras AC, Raught B, Sonenberg N. Regulation of translation initiation by FRAP/mTOR. Genes Dev. 2001;15:807–826. - PubMed
-
- Komar AA, Hatzoglou M. Internal ribosome entry sites in cellular mRNAs: Mystery of their existence. J Biol Chem. 2005;280:23425–23428. - PubMed
-
- Shaw RJ, Cantley LC. Ras, PI(3)K and mTOR signaling controls tumour cell growth. Nature. 2006;441:424–30. - PubMed
-
- Brunn GJ, et al. Phosphorylation of the translational repressor PHAS-I by the mammalian target of rapamycin. Science. 1997;277:99–101. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
