

Conformation and membrane position of the region linking the two C2 domains in synaptotagmin 1 by site-directed spin labeling
- PMID: 18956883
- PMCID: PMC2661311
- DOI: 10.1021/bi801470m
Conformation and membrane position of the region linking the two C2 domains in synaptotagmin 1 by site-directed spin labeling
Abstract
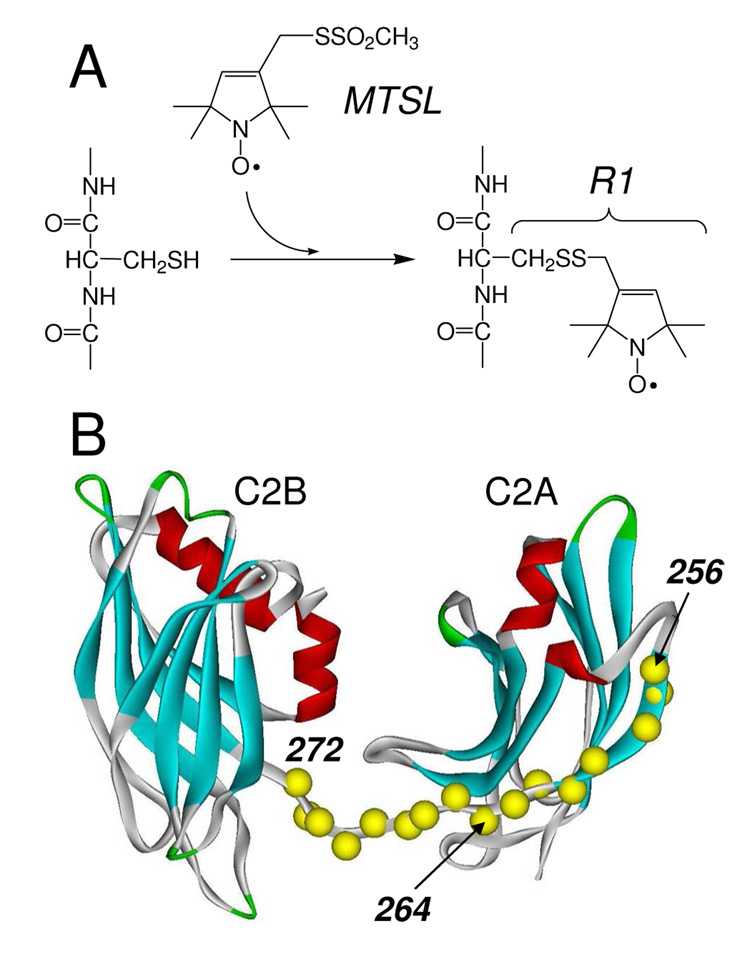
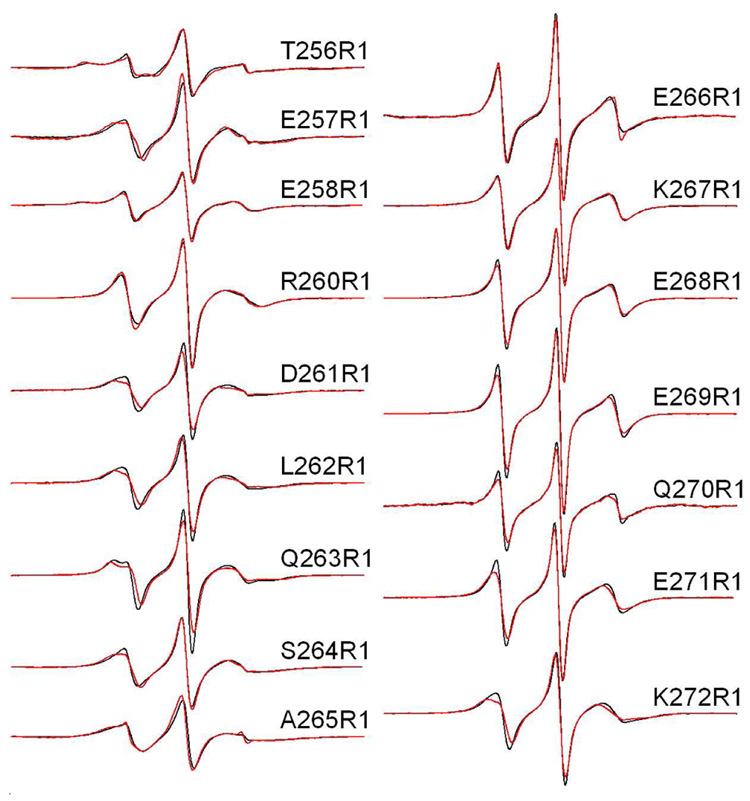
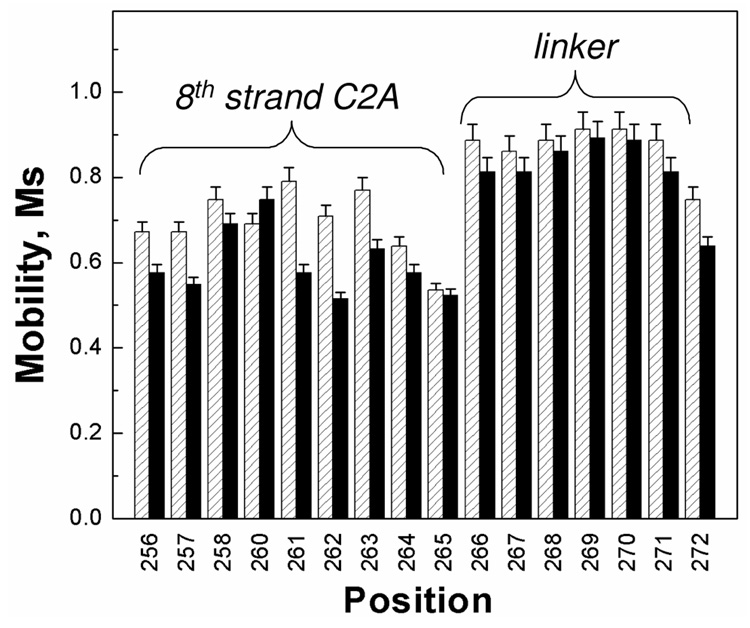
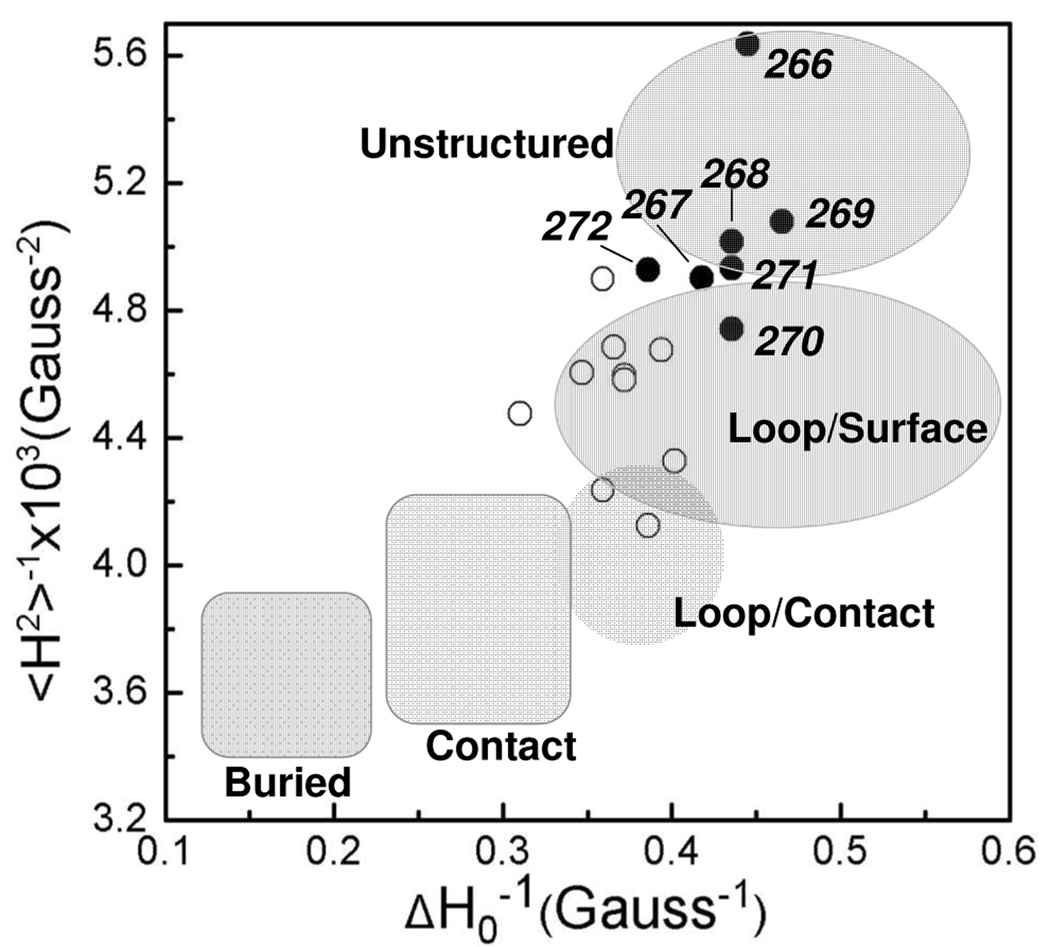
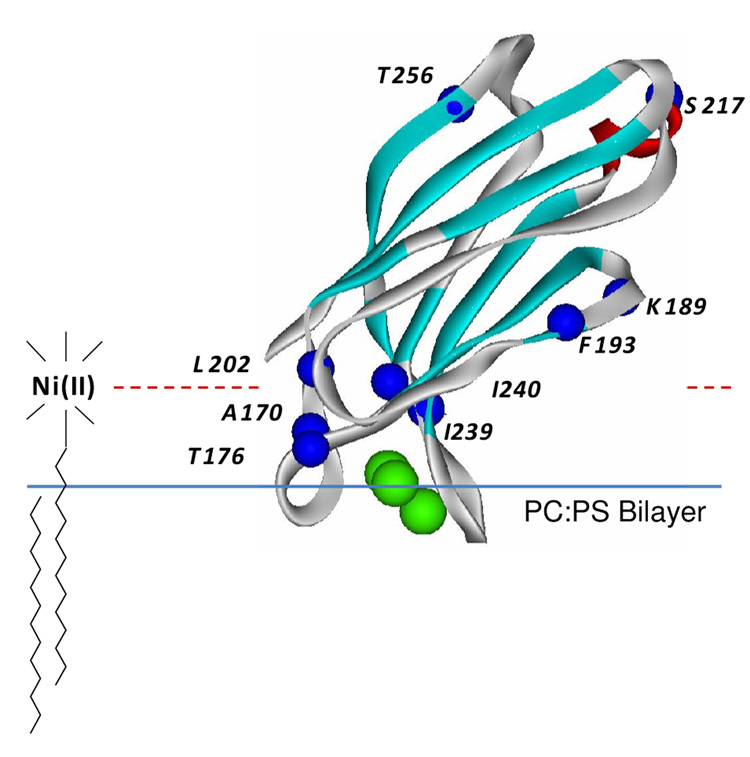
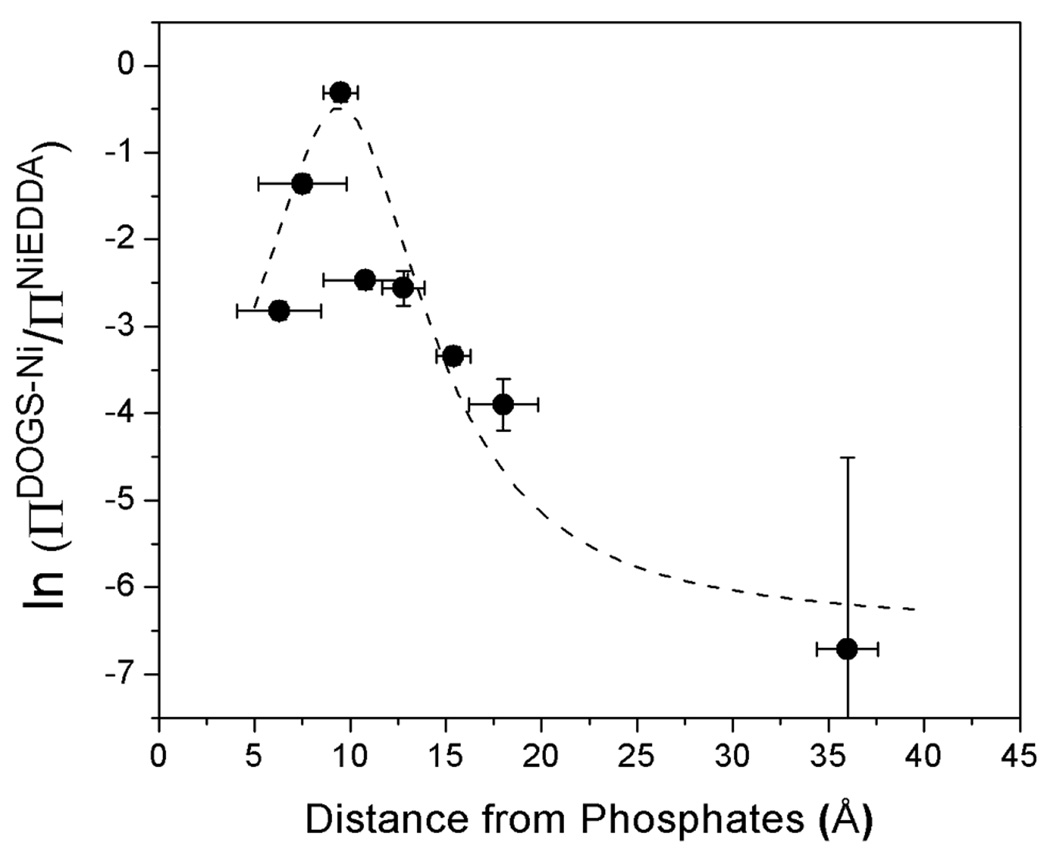
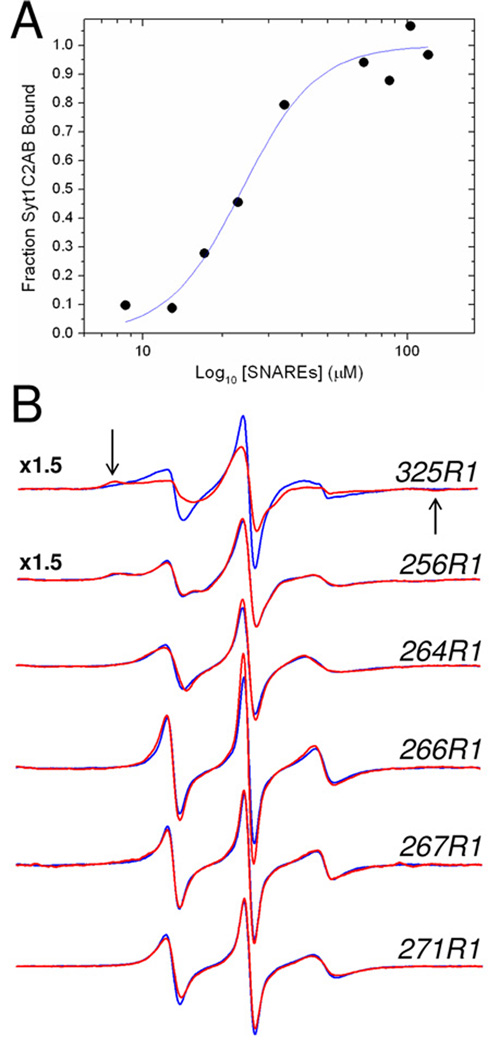
Synaptotagmin 1 (syt1) is an integral membrane protein localized on the synaptic vesicle that acts as the Ca(2+) sensor for neuronal exocytosis. Synaptotagmin 1 contains two C2 domains, C2A and C2B, which bind Ca(2+) ions, membranes, and SNAREs. Here, site-directed spin labeling (SDSL) was used to determine the position and dynamics of the region that links the two C2 domains in a water soluble construct encompassing the two C2 domains (syt1C2AB). An analysis of the EPR line shapes from this region indicates that the linker is flexible and unstructured when syt1 is in solution or bound to lipid bilayers. The nanosecond dynamics of the linker does not change, in the presence or absence of Ca(2+), suggesting that there is no Ca(2+)-dependent intramolecular association between the two domains. When syt1C2AB is membrane-bound, the position of the linker relative to the membrane interface was determined by measuring parameters for the collision of the spin-labeled syt1C2AB mutants with both soluble and membrane-bound Ni(II) chelates. These data indicate that the linker does not penetrate the membrane surface but lies approximately 7-10 A from the bilayer surface. In addition, the linker remains flexible when syt1C2AB binds to the SNARE complex, indicating that direct interactions between this linker and the SNAREs do not mediate association. These data suggest that the two C2 domains of syt1 interact independently on the membrane interface, or when bound to SNAREs.
Figures
References
-
- Jahn R, Lang T, Suhof TC. Membrane Fusion. Cell. 2003;112:519–533. - PubMed
-
- Rothman JE. Mechanisms of intracellular protein transport. Nature. 1994;372:55–63. - PubMed
-
- Sudhof TC. The synaptic vesicle cycle. Annual Review of Neuroscience. 2004;27:509–547. - PubMed
-
- Koh TW, Bellen HJ. Synaptotagmin I, a Ca2+ sensor for neurotransmitter release. Trends in Neurosciences. 2003;26:413–422. - PubMed
-
- Rizo J, Chen X, Arac D. Unraveling the mechanisms of synaptotagmin and SNARE function in neurotransmitter release. Trends Cell Biol. 2006;16:339–350. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
