

Fibrils connect microtubule tips with kinetochores: a mechanism to couple tubulin dynamics to chromosome motion
- PMID: 18957206
- PMCID: PMC2746696
- DOI: 10.1016/j.cell.2008.08.038
Fibrils connect microtubule tips with kinetochores: a mechanism to couple tubulin dynamics to chromosome motion
Abstract
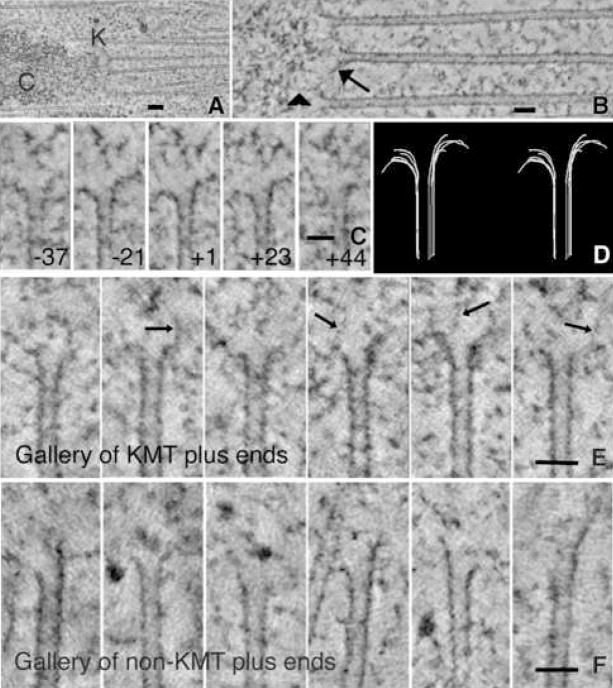
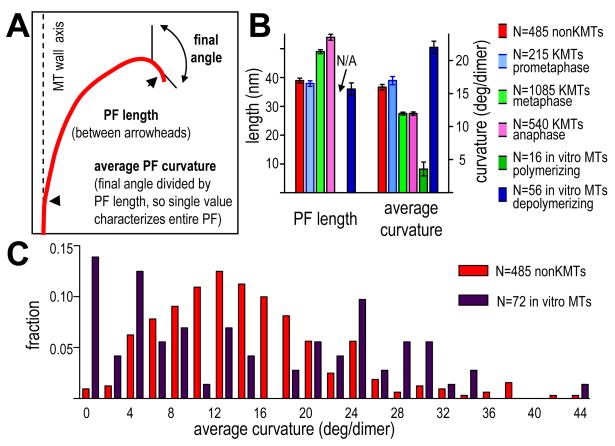
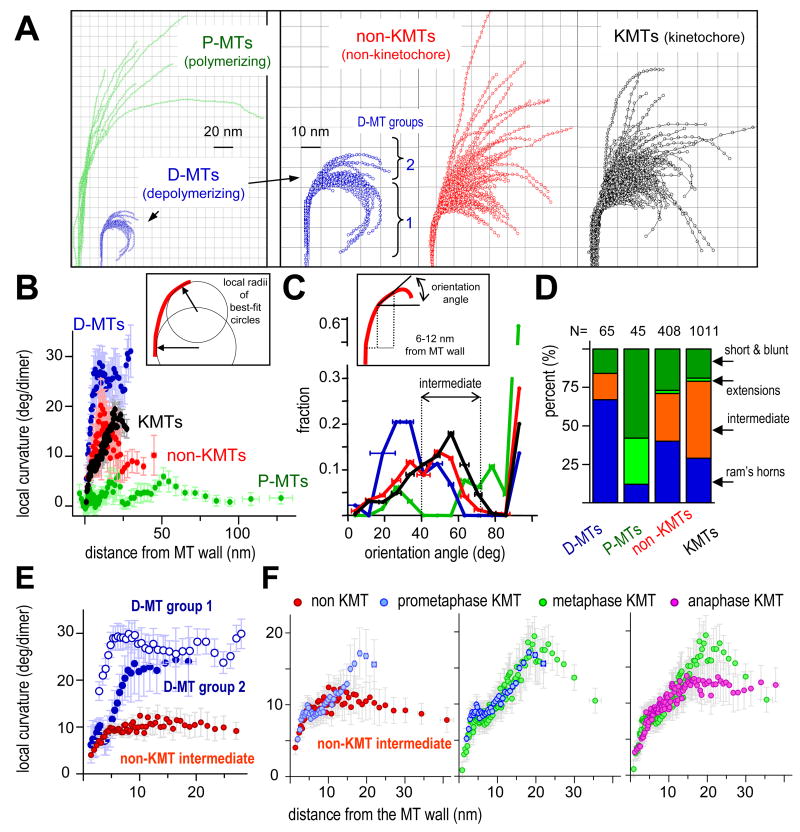
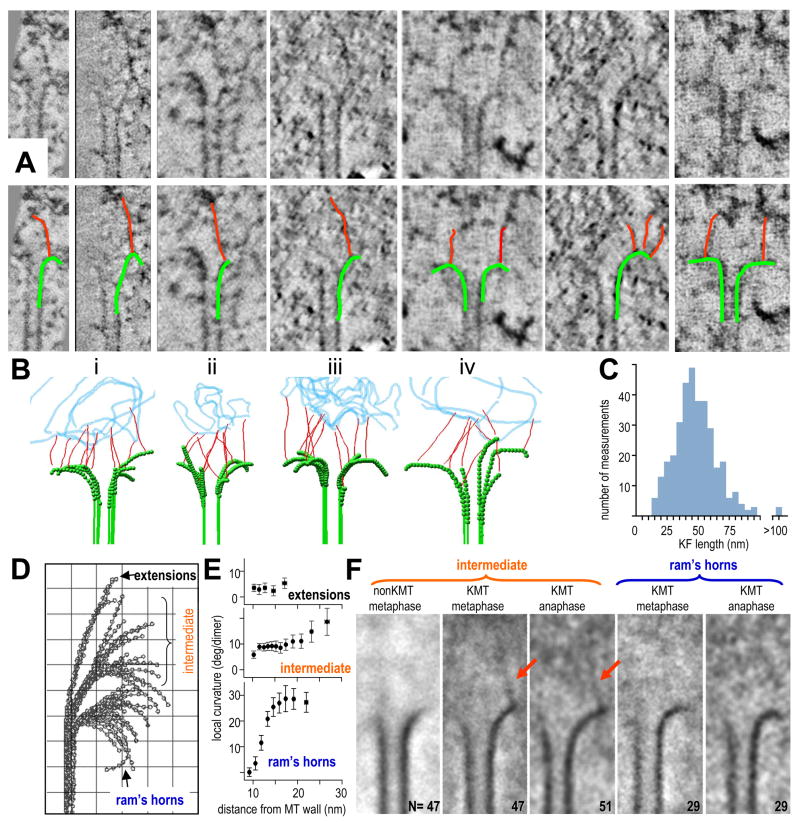
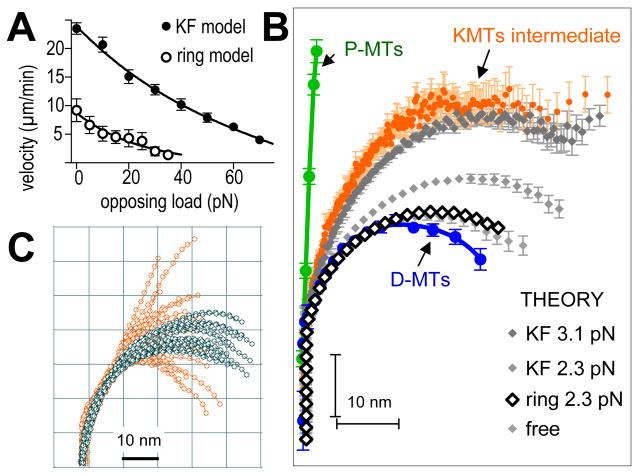
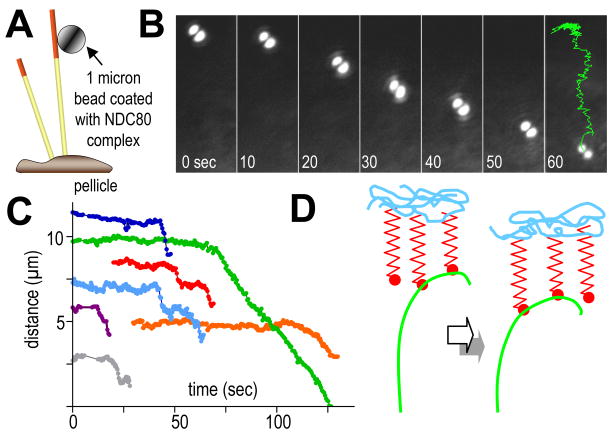
Kinetochores of mitotic chromosomes are coupled to spindle microtubules in ways that allow the energy from tubulin dynamics to drive chromosome motion. Most kinetochore-associated microtubule ends display curving "protofilaments," strands of tubulin dimers that bend away from the microtubule axis. Both a kinetochore "plate" and an encircling, ring-shaped protein complex have been proposed to link protofilament bending to poleward chromosome motion. Here we show by electron tomography that slender fibrils connect curved protofilaments directly to the inner kinetochore. Fibril-protofilament associations correlate with a local straightening of the flared protofilaments. Theoretical analysis reveals that protofilament-fibril connections would be efficient couplers for chromosome motion, and experimental work on two very different kinetochore components suggests that filamentous proteins can couple shortening microtubules to cargo movements. These analyses define a ring-independent mechanism for harnessing microtubule dynamics directly to chromosome movement.
Figures
Comment in
-
Kinetochores and microtubules wed without a ring.Cell. 2008 Oct 17;135(2):211-3. doi: 10.1016/j.cell.2008.10.001. Cell. 2008. PMID: 18957196 Free PMC article.
References
-
- Arnal I, Heichette C, Diamantopoulos GS, Chretien D. CLIP-170/tubulin-curved oligomers coassemble at microtubule ends and promote rescues. Curr Biol. 2004;14:2086–2095. - PubMed
-
- Cheeseman IM, Chappie JS, Wilson-Kubalek EM, Desai A. The conserved KMN network constitutes the core microtubule-binding site of the kinetochore. Cell. 2006;127:983–997. - PubMed
-
- Cheeseman IM, Desai A. Molecular architecture of the kinetochore-microtubule interface. Nat Rev Mol Cell Biol. 2008;9:33–46. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
