

Cystatin C-cathepsin B axis regulates amyloid beta levels and associated neuronal deficits in an animal model of Alzheimer's disease
- PMID: 18957217
- PMCID: PMC2755563
- DOI: 10.1016/j.neuron.2008.10.001
Cystatin C-cathepsin B axis regulates amyloid beta levels and associated neuronal deficits in an animal model of Alzheimer's disease
Abstract
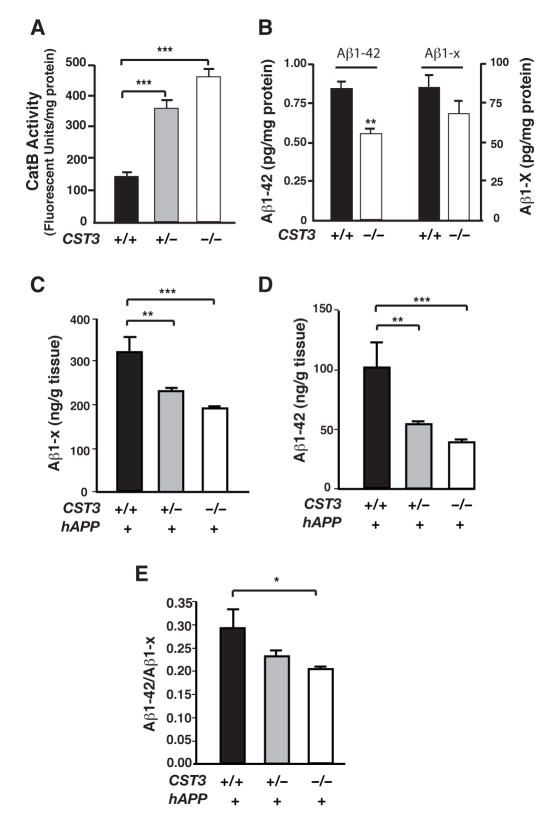
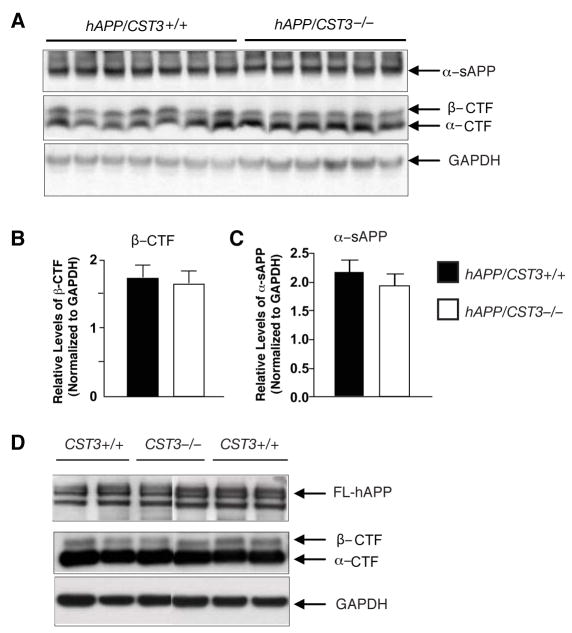
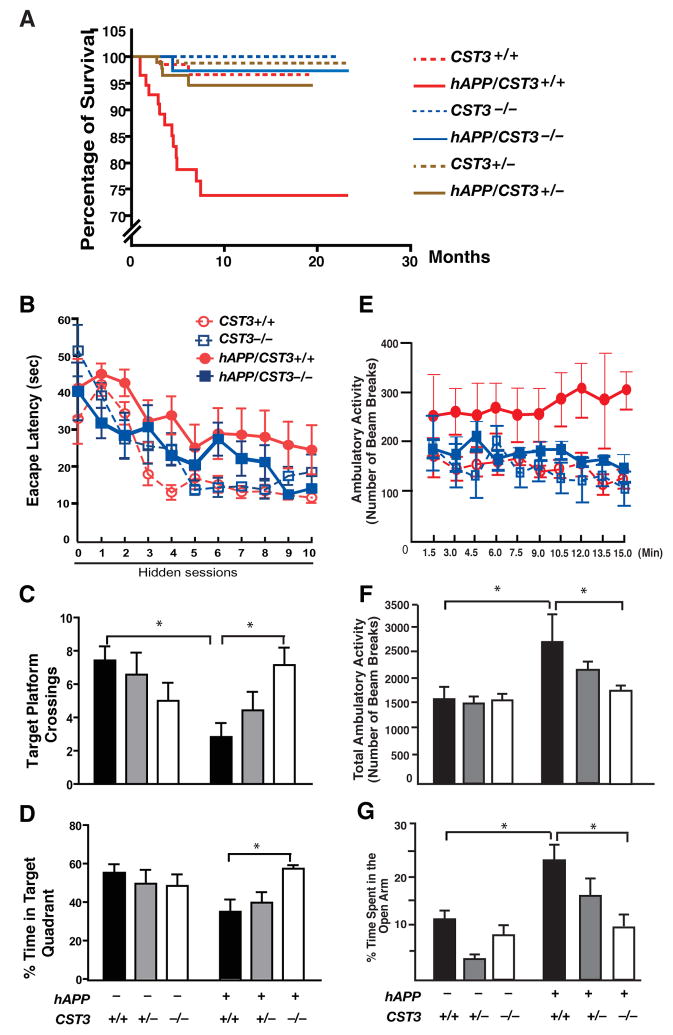
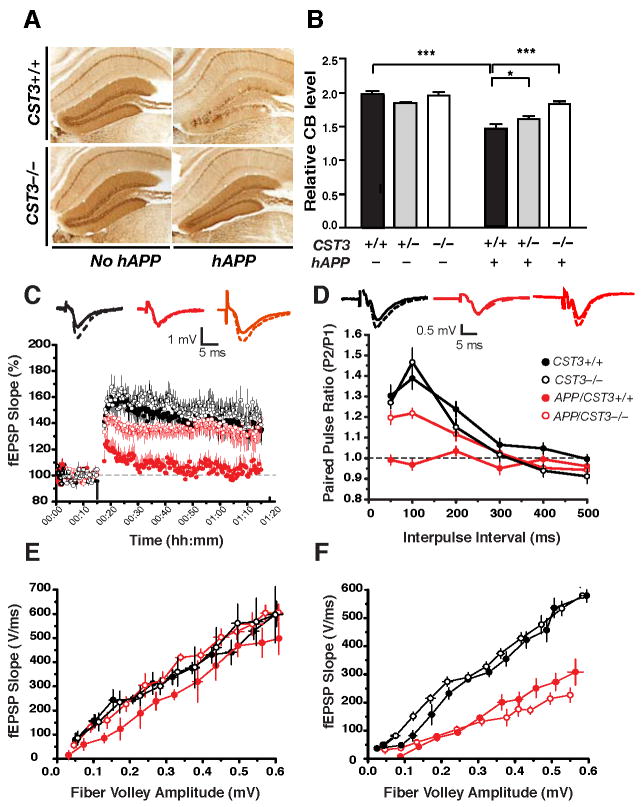
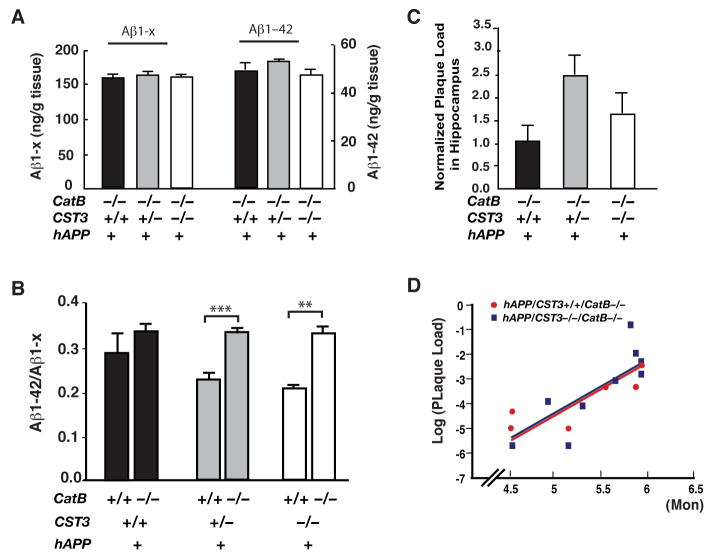
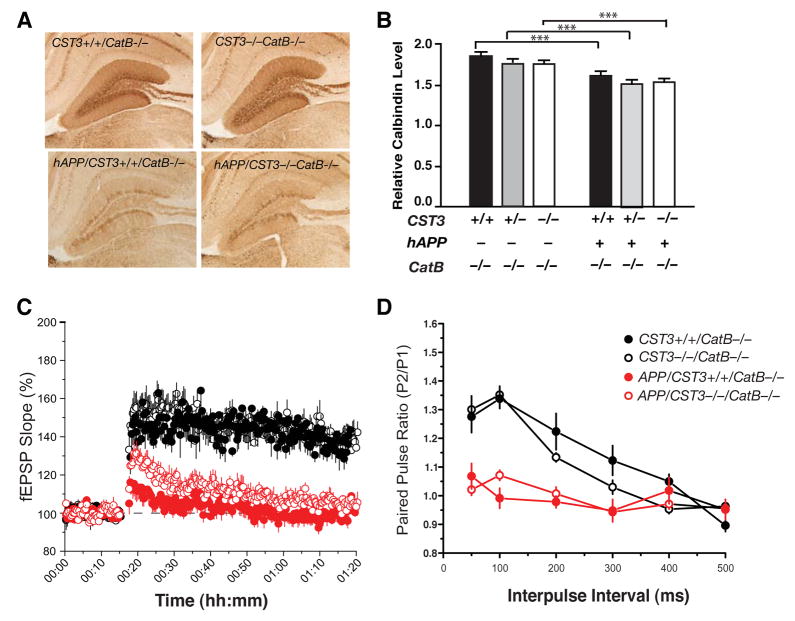
Impaired degradation of amyloid beta (Abeta) peptides could lead to Abeta accumulation, an early trigger of Alzheimer's disease (AD). How Abeta-degrading enzymes are regulated remains largely unknown. Cystatin C (CysC, CST3) is an endogenous inhibitor of cysteine proteases, including cathepsin B (CatB), a recently discovered Abeta-degrading enzyme. A CST3 polymorphism is associated with an increased risk of late-onset sporadic AD. Here, we identified CysC as the key inhibitor of CatB-induced Abeta degradation in vivo. Genetic ablation of CST3 in hAPP-J20 mice significantly lowered soluble Abeta levels, the relative abundance of Abeta1-42, and plaque load. CysC removal also attenuated Abeta-associated cognitive deficits and behavioral abnormalities and restored synaptic plasticity in the hippocampus. Importantly, the beneficial effects of CysC reduction were abolished on a CatB null background, providing direct evidence that CysC regulates soluble Abeta and Abeta-associated neuronal deficits through inhibiting CatB-induced Abeta degradation.
Figures

References
-
- Abrahamson M, Barrett AJ, Salvesen G, Grubb A. Isolation of six cysteine proteinase inhibitors from human urine. Their physicochemical and enzyme kinetic properties and concentrations in biological fluids. J Biol Chem. 1986;261:11282–11289. - PubMed
-
- Benussi L, Ghidoni R, Steinhoff T, Alberici A, Villa A, Mazzoli F, Nicosia F, Barbiero L, Broglio L, Feudatari E, et al. Alzheimer disease-associated cystatin C variant undergoes impaired secretion. Neurobiol Dis. 2003;13:15–21. - PubMed
-
- Bertram L, McQueen MB, Mullin K, Blacker D, Tanzi RE. Systematic meta-analyses of Alzheimer disease genetic association studies: the AlzGene database. Nat Genet. 2007;39:17–23. - PubMed
-
- Beyer K, Lao JI, Gomez M, Riutort N, Latorre P, Mate JL, Ariza A. Alzheimer’s disease and the cystatin C gene polymorphism: An association study. Neurosci Lett. 2001;315:17–20. - PubMed
-
- Bogyo M, Verhelst S, Bellingard-Dubouchaud V, Toba S, Greenbaum D. Selective targeting of lysosomal cysteine proteases with radiolabeled electrophilic substrate analogs. Chem Biol. 2000;7:27–38. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
