

Proton-coupled protein transport through the anthrax toxin channel
- PMID: 18957378
- PMCID: PMC2674091
- DOI: 10.1098/rstb.2008.0126
Proton-coupled protein transport through the anthrax toxin channel
Abstract
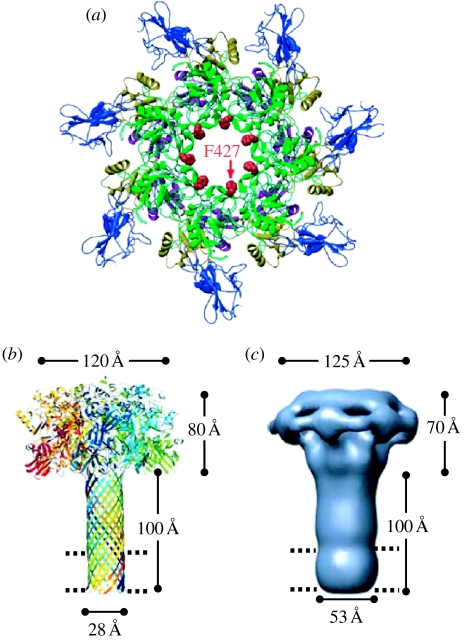
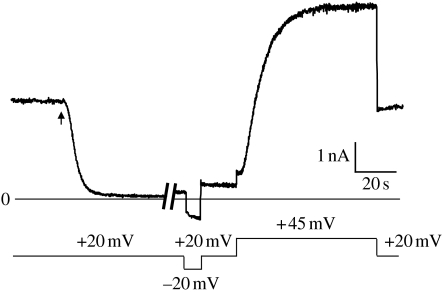
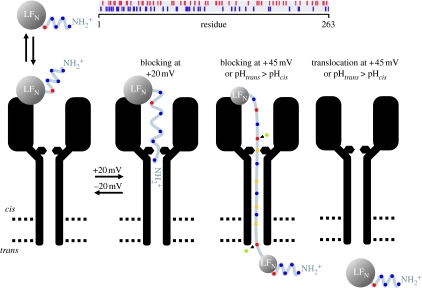
Anthrax toxin consists of three proteins (approx. 90kDa each): lethal factor (LF); oedema factor (OF); and protective antigen (PA). The former two are enzymes that act when they reach the cytosol of a targeted cell. To enter the cytosol, however, which they do after being endocytosed into an acidic vesicle compartment, they require the third component, PA. PA (or rather its proteolytically generated fragment PA63) forms at low pH a heptameric beta-barrel channel, (PA63)7, through which LF and OF are transported--a phenomenon we have demonstrated in planar phospholipid bilayers. It might appear that (PA63)7 simply forms a large hole through which LF and OF diffuse. However, LF and OF are folded proteins, much too large to fit through the approximately 15A diameter (PA63)7 beta-barrel. This paper discusses how the (PA63)7 channel both participates in the unfolding of LF and OF and functions in their translocation as a proton-protein symporter.
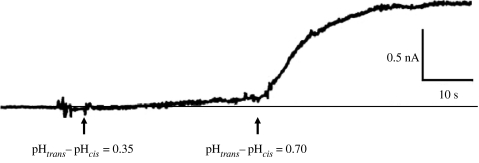
Figures
References
-
- Blaustein R.O., Finkelstein A. Voltage-dependent block of anthrax toxin channels in planar phospholipid bilayer membranes by symmetric tetraalkylammonium ions: effects on macroscopic conductance. J. Gen. Physiol. 1990a;96:905–919. doi:10.1085/jgp.96.5.905 - DOI - PMC - PubMed
-
- Blaustein R.O., Finkelstein A. Diffusion limitation in the block by symmetric tetraalkylammonium ions of anthrax toxin channels in planar phospholipid bilayer membranes. J. Gen. Physiol. 1990b;96:943–957. doi:10.1085/jgp.96.5.943 - DOI - PMC - PubMed
-
- Falnes P.O., Sandvig K. Penetration of protein toxins into cells. Curr. Opin. Cell Biol. 2000;12:407–413. doi:10.1016/S0955-0674(00)00109-5 - DOI - PubMed
-
- Falnes P.O., Madshas I.H., Sandvig K., Olsnes S. Replacement of negative by positive charges in the presumed membrane-inserted part of diphtheria toxin B fragment. Effect on membrane translocation and on formation of cation channels. J. Biol. Chem. 1992;267:12 284–12 290. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials