

Neural substrate of concurrent sound perception: direct electrophysiological recordings from human auditory cortex
- PMID: 18958219
- PMCID: PMC2525982
- DOI: 10.3389/neuro.09.005.2007
Neural substrate of concurrent sound perception: direct electrophysiological recordings from human auditory cortex
Abstract
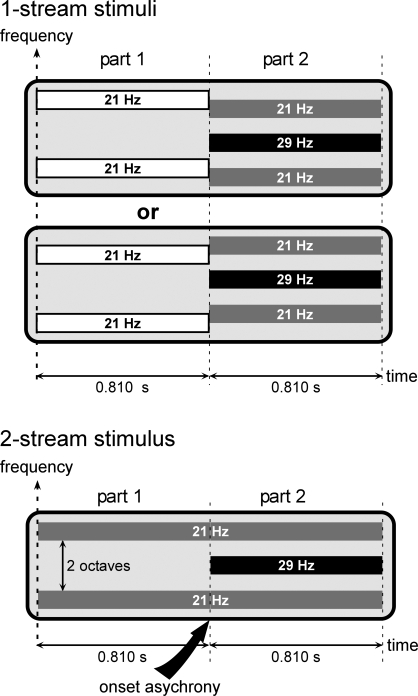
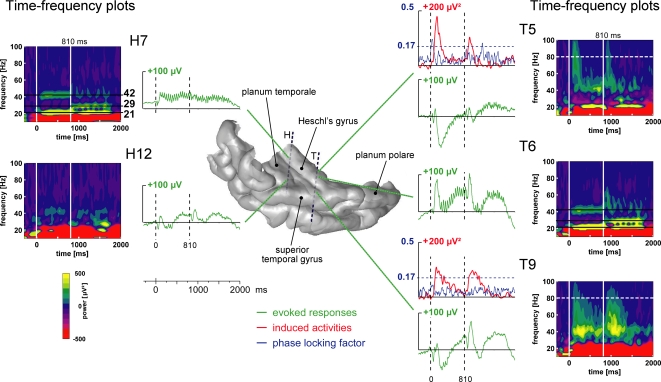
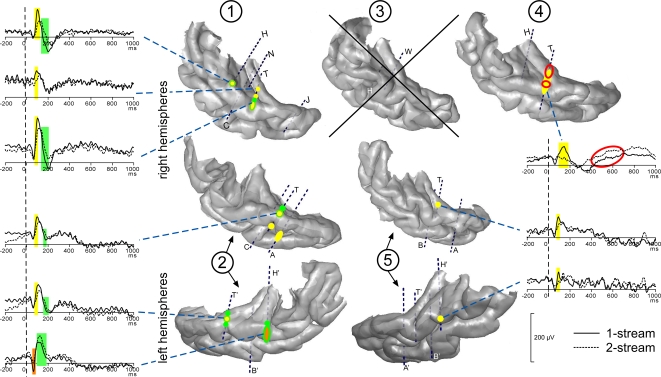
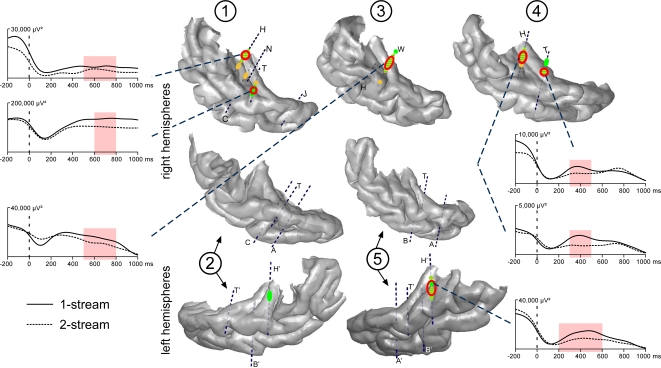
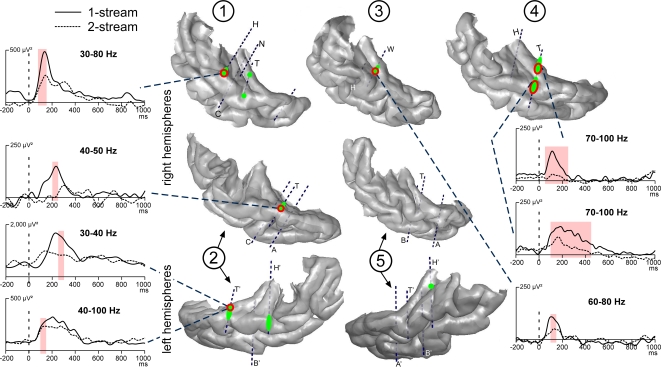
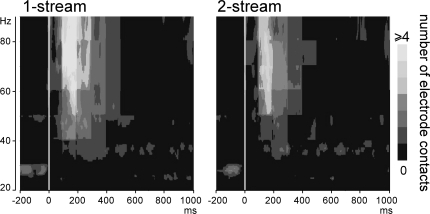
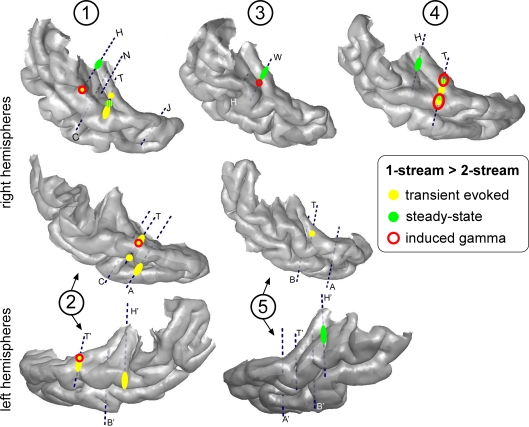
In everyday life, consciously or not, we are constantly disentangling the multiple auditory sources contributing to our acoustical environment. To better understand the neural mechanisms involved in concurrent sound processing, we manipulated sound onset asynchrony to induce the segregation or grouping of two concurrent sounds. Each sound consisted of amplitude-modulated tones at different carrier and modulation frequencies, allowing a cortical tagging of each sound. Electrophysiological recordings were carried out in epileptic patients with pharmacologically resistant partial epilepsy, implanted with depth electrodes in the temporal cortex. Patients were presented with the stimuli while they performed an auditory distracting task. We found that transient and steady-state evoked responses, and induced gamma oscillatory activities were enhanced in the case of onset synchrony. These effects were mainly located in the Heschl's gyrus for steady-state responses whereas they were found in the lateral superior temporal gyrus for evoked transient responses and induced gamma oscillations. They can be related to distinct neural mechanisms such as frequency selectivity and habituation. These results in the auditory cortex provide an anatomically refined description of the neurophysiological components which might be involved in the perception of concurrent sounds.
Keywords: EEG; auditory cortex; event-related; gamma; human; steady-state; stream segregation.
Figures
Similar articles
-
Effects of selective attention on the electrophysiological representation of concurrent sounds in the human auditory cortex.J Neurosci. 2007 Aug 29;27(35):9252-61. doi: 10.1523/JNEUROSCI.1402-07.2007. J Neurosci. 2007. PMID: 17728439 Free PMC article.
-
Neural processes of auditory perception in Heschl's gyrus for upcoming acoustic stimuli in humans.Hear Res. 2020 Mar 15;388:107895. doi: 10.1016/j.heares.2020.107895. Epub 2020 Jan 18. Hear Res. 2020. PMID: 31982643
-
Consonance and dissonance of musical chords: neural correlates in auditory cortex of monkeys and humans.J Neurophysiol. 2001 Dec;86(6):2761-88. doi: 10.1152/jn.2001.86.6.2761. J Neurophysiol. 2001. PMID: 11731536 Clinical Trial.
-
Auditory processing in the human cortex: An intracranial electrophysiology perspective.Laryngoscope Investig Otolaryngol. 2017 Apr 12;2(4):147-156. doi: 10.1002/lio2.73. eCollection 2017 Aug. Laryngoscope Investig Otolaryngol. 2017. PMID: 28894834 Free PMC article. Review.
-
[Auditory perception and language: functional imaging of speech sensitive auditory cortex].Rev Neurol (Paris). 2001 Sep;157(8-9 Pt 1):837-46. Rev Neurol (Paris). 2001. PMID: 11677406 Review. French.
Cited by
-
Cortical gamma-oscillations modulated by auditory-motor tasks-intracranial recording in patients with epilepsy.Hum Brain Mapp. 2010 Nov;31(11):1627-42. doi: 10.1002/hbm.20963. Hum Brain Mapp. 2010. PMID: 20143383 Free PMC article.
-
Across-subjects classification of stimulus modality from human MEG high frequency activity.PLoS Comput Biol. 2018 Mar 12;14(3):e1005938. doi: 10.1371/journal.pcbi.1005938. eCollection 2018 Mar. PLoS Comput Biol. 2018. PMID: 29529062 Free PMC article.
-
Neurophysiological mechanisms involved in auditory perceptual organization.Front Neurosci. 2009 Sep 15;3(2):182-91. doi: 10.3389/neuro.01.025.2009. eCollection 2009 Sep. Front Neurosci. 2009. PMID: 20011140 Free PMC article.
-
EEG signatures accompanying auditory figure-ground segregation.Neuroimage. 2016 Nov 1;141:108-119. doi: 10.1016/j.neuroimage.2016.07.028. Epub 2016 Jul 12. Neuroimage. 2016. PMID: 27421185 Free PMC article.
-
Neural mechanisms of rhythmic masking release in monkey primary auditory cortex: implications for models of auditory scene analysis.J Neurophysiol. 2012 May;107(9):2366-82. doi: 10.1152/jn.01010.2011. Epub 2012 Feb 8. J Neurophysiol. 2012. PMID: 22323627 Free PMC article.
References
LinkOut - more resources
Full Text Sources