

Developmental sensitivity of hippocampal interneurons to ethanol: involvement of the hyperpolarization-activated current, Ih
- PMID: 18971298
- PMCID: PMC2637011
- DOI: 10.1152/jn.90557.2008
Developmental sensitivity of hippocampal interneurons to ethanol: involvement of the hyperpolarization-activated current, Ih
Abstract
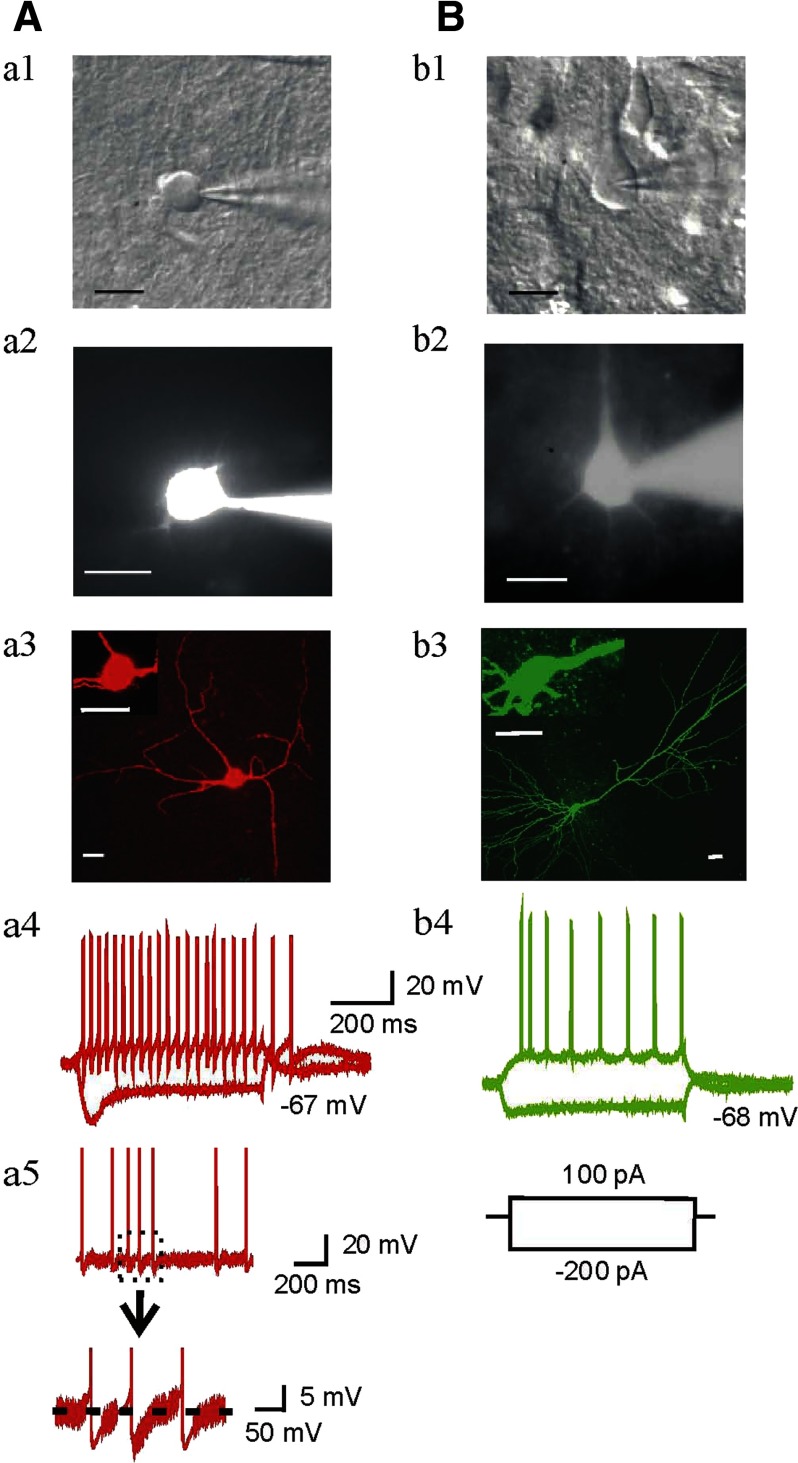
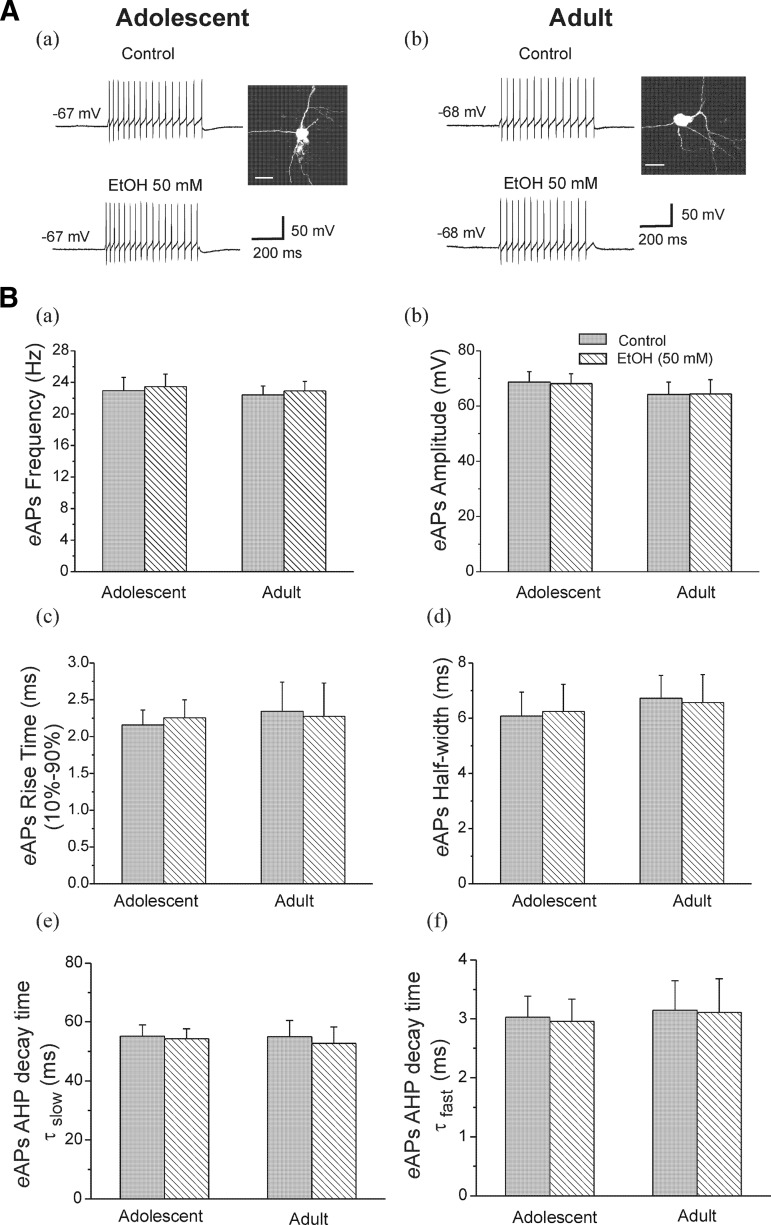
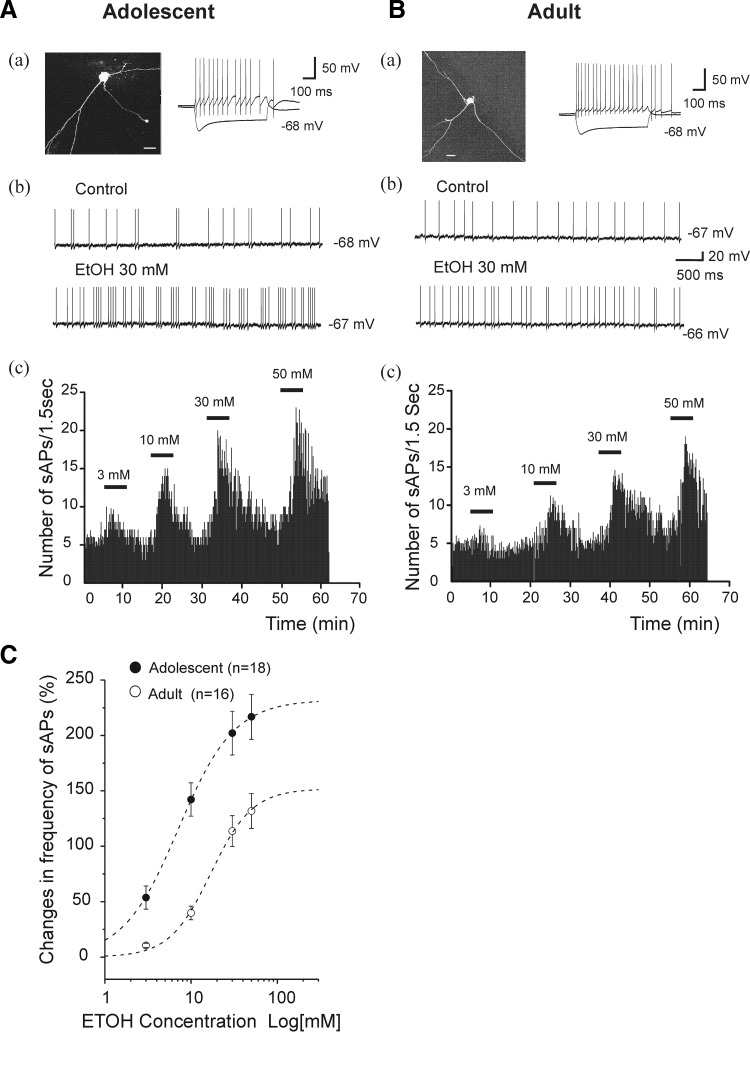
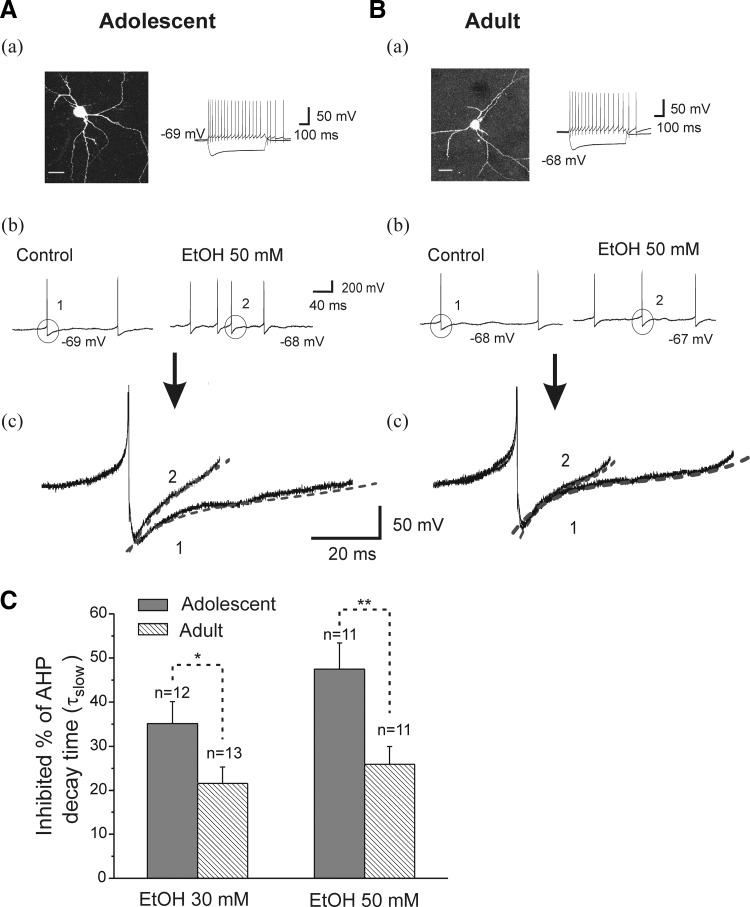
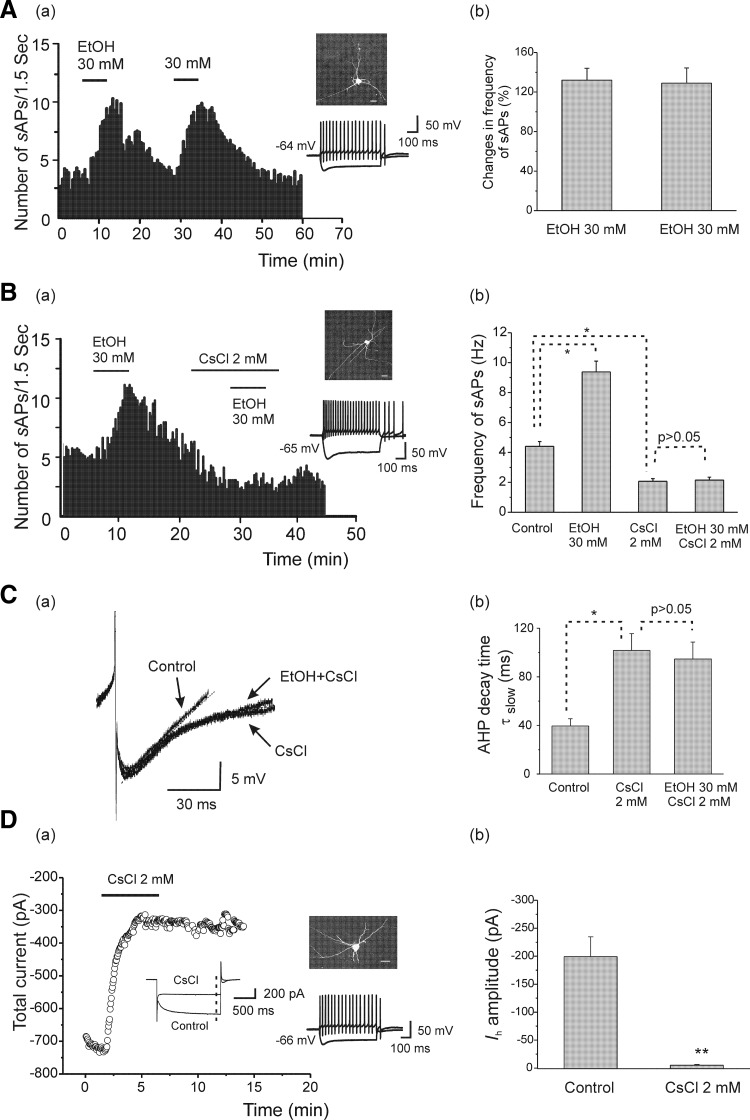
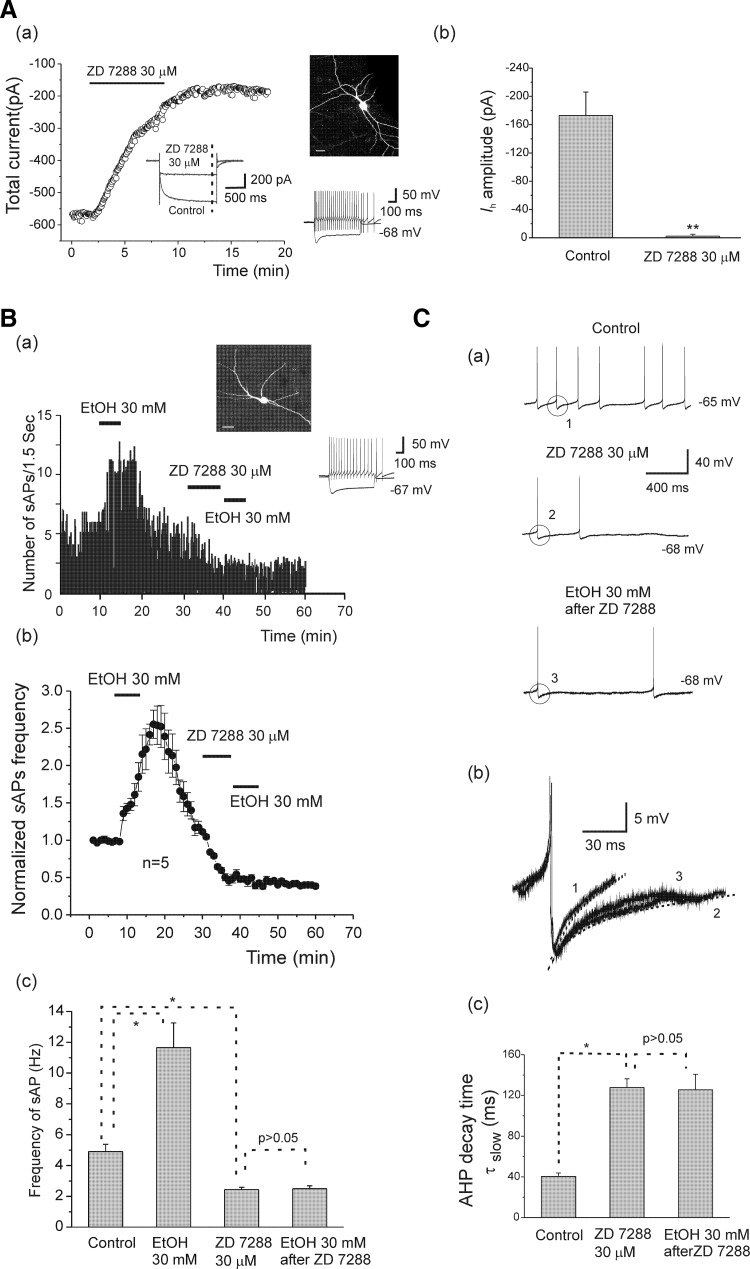
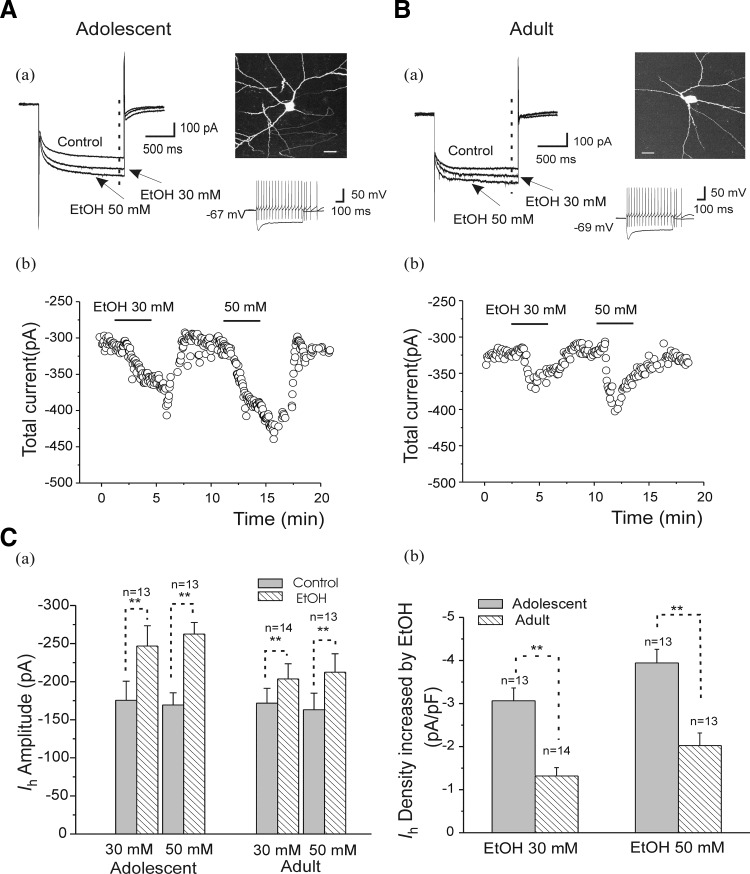
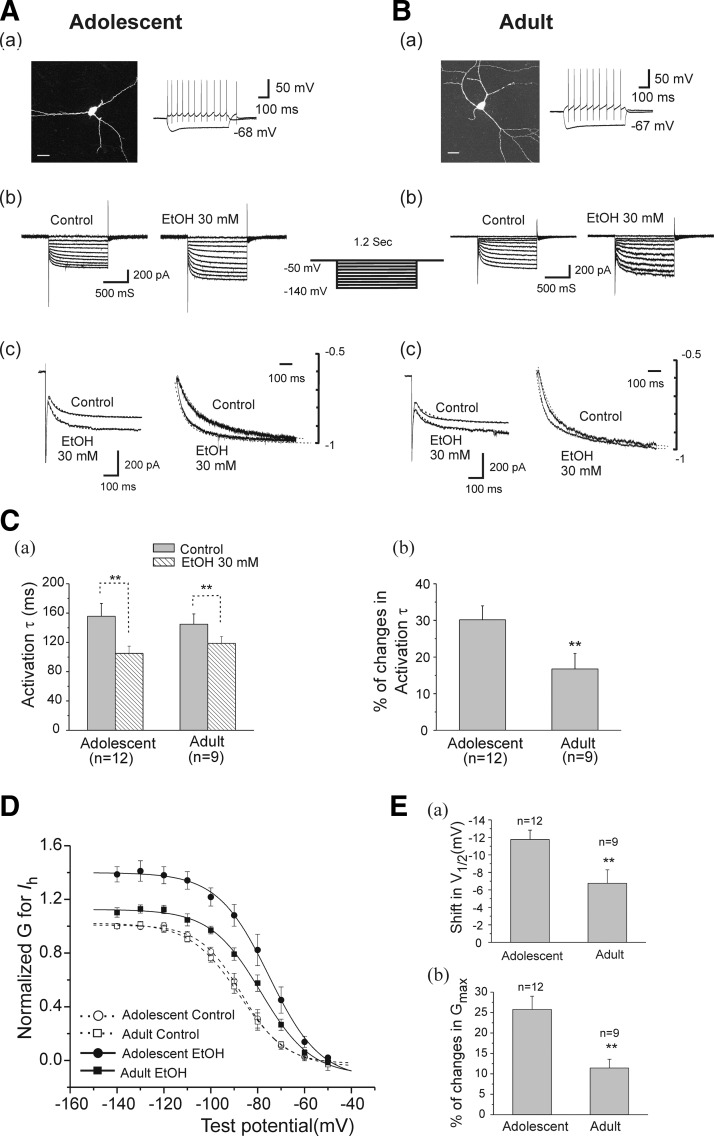
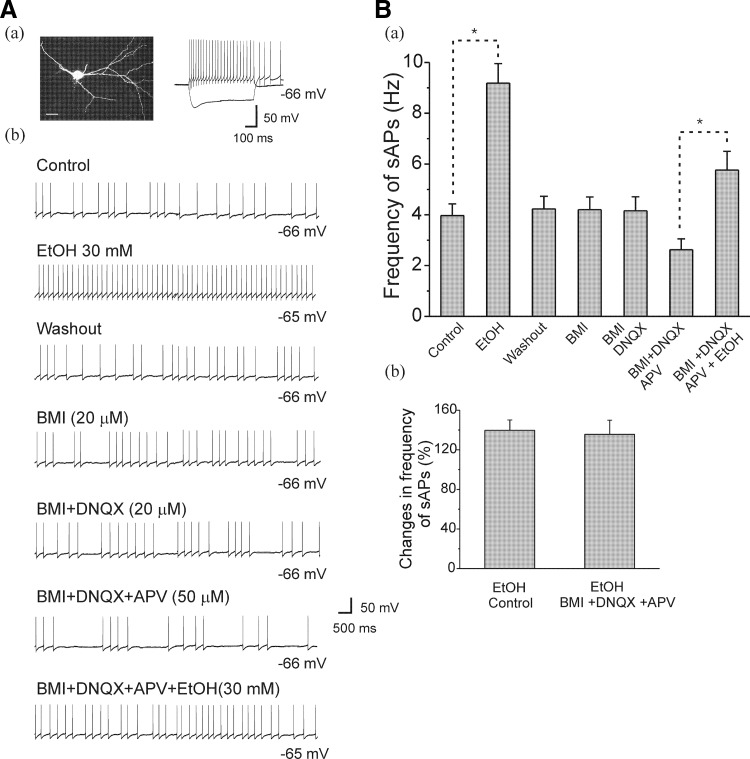
Ethanol (EtOH) has powerful effects on GABA(A) receptor-mediated neurotransmission, and we have previously shown that EtOH-induced enhancement of GABA(A) receptor-mediated synaptic transmission in the hippocampus is developmentally regulated. Because synaptic inhibition is determined in part by the firing properties of interneurons, we have investigated the mechanisms whereby EtOH influences the spontaneous firing characteristics and hyperpolarization-activated cation current (Ih) of hippocampal interneurons located in the near to the border of stratum lacunosum moleculare and s. radiatum of adolescent and adult rats. EtOH did not affect current injection-induced action potentials of interneurons that do not exhibit spontaneous firing. However, in neurons that fire spontaneously, EtOH enhanced the frequency of spontaneous action potentials (sAPs) in a concentration-dependent manner, an effect that was more pronounced in interneurons from adolescent rats, compared with adult rats. EtOH also modulated the afterhyperpolarization (AHP) that follows sAPs by shortening the tau(slow) decay time constant, and this effect was more pronounced in slices from adolescent rats. EtOH increased Ih amplitudes, accelerated Ih activation kinetics, and increased the maximal Ih conductance in interneurons from animals in both age groups. These effects were also more pronounced in interneurons from adolescents and persisted in the presence of glutamatergic and GABAergic blockers. However, EtOH failed to affect sAP firing in the presence of ZD7288 or cesium chloride. These results suggest that Ih may be of mechanistic significance in the effect of EtOH on interneuron spontaneous firing.
Figures
Similar articles
-
Excitation of rat cerebellar Golgi cells by ethanol: further characterization of the mechanism.Alcohol Clin Exp Res. 2012 Apr;36(4):616-24. doi: 10.1111/j.1530-0277.2011.01658.x. Epub 2011 Oct 17. Alcohol Clin Exp Res. 2012. PMID: 22004123 Free PMC article.
-
Differential sensitivity of hippocampal interneurons to ethanol in adolescent and adult rats.J Pharmacol Exp Ther. 2010 Oct;335(1):51-60. doi: 10.1124/jpet.110.168450. Epub 2010 Jul 21. J Pharmacol Exp Ther. 2010. PMID: 20660126 Free PMC article.
-
Bi-directional modulation of hyperpolarization-activated cation currents (Ih) by ethanol in rat hippocampal CA3 pyramidal neurons.Neuropharmacology. 2023 Apr 1;227:109423. doi: 10.1016/j.neuropharm.2023.109423. Epub 2023 Jan 20. Neuropharmacology. 2023. PMID: 36690323
-
Changes in the Properties of Ethanol-Sensitive Molecular Targets During Maturation and Aging.Adv Exp Med Biol. 2025;1473:299-316. doi: 10.1007/978-3-031-81908-7_13. Adv Exp Med Biol. 2025. PMID: 40128484 Review.
-
[Cesium].Encephale. 1979;5(4):359-74. Encephale. 1979. PMID: 230026 Review. French.
Cited by
-
Simulation of alcohol action upon a detailed Purkinje neuron model and a simpler surrogate model that runs >400 times faster.BMC Neurosci. 2015 Apr 26;16:27. doi: 10.1186/s12868-015-0162-6. BMC Neurosci. 2015. PMID: 25928094 Free PMC article.
-
Excitation of rat cerebellar Golgi cells by ethanol: further characterization of the mechanism.Alcohol Clin Exp Res. 2012 Apr;36(4):616-24. doi: 10.1111/j.1530-0277.2011.01658.x. Epub 2011 Oct 17. Alcohol Clin Exp Res. 2012. PMID: 22004123 Free PMC article.
-
Alcohol excites cerebellar Golgi cells by inhibiting the Na+/K+ ATPase.Neuropsychopharmacology. 2010 Aug;35(9):1984-96. doi: 10.1038/npp.2010.76. Epub 2010 Jun 2. Neuropsychopharmacology. 2010. PMID: 20520600 Free PMC article.
-
Adolescent ethanol exposure: does it produce long-lasting electrophysiological effects?Alcohol. 2010 Feb;44(1):27-37. doi: 10.1016/j.alcohol.2009.09.033. Alcohol. 2010. PMID: 20113872 Free PMC article. Review.
-
Regulation of epileptiform discharges in rat neocortex by HCN channels.J Neurophysiol. 2013 Oct;110(8):1733-43. doi: 10.1152/jn.00955.2012. Epub 2013 Jul 17. J Neurophysiol. 2013. PMID: 23864381 Free PMC article.
References
-
- Allansson L, Khatibi S, Olsson T, Hansson E. Acute ethanol exposure induces transients, cell swelling and transformation of actin cytoskeleton in astroglial primary cultures. J Neurochem 76: 472–479, 2001. - PubMed
-
- Appel SB, Liu Z, McElvain MA, Brodie MS. Ethanol excitation of dopaminergic ventral tegmental area neurons is blocked by quinidine. J Pharmacol Exp Ther 306: 437–446, 2003. - PubMed
-
- Bender RA, Brewster A, Santoro B, Ludwig A, Hofmann F, Biel M, Baram TZ. Differential and age-dependent expression of hyperpolarization-activated, cyclic nucleotide-gated cation channel isoforms 1-4 suggests evolving roles in the developing rat hippocampus. Neuroscience 106: 689–698, 2001. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous