

Aldehyde dehydrogenase 1a1 is dispensable for stem cell function in the mouse hematopoietic and nervous systems
- PMID: 18971422
- PMCID: PMC2647681
- DOI: 10.1182/blood-2008-05-156752
Aldehyde dehydrogenase 1a1 is dispensable for stem cell function in the mouse hematopoietic and nervous systems
Abstract
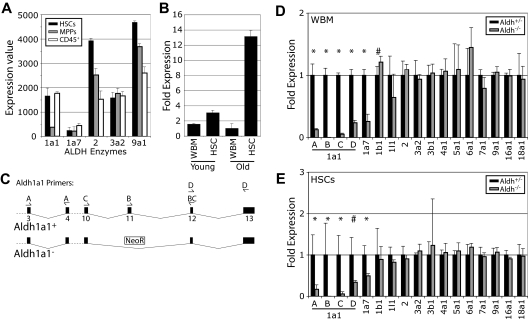
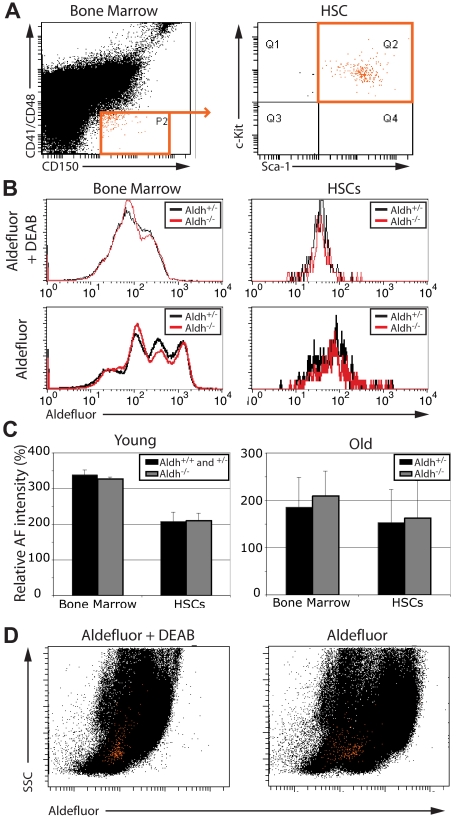
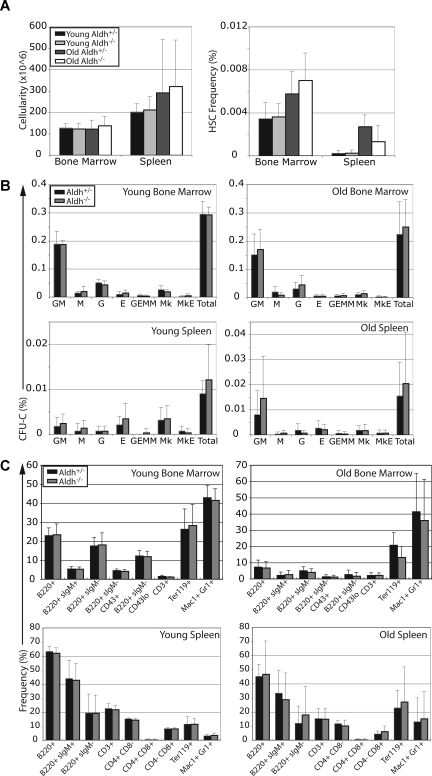
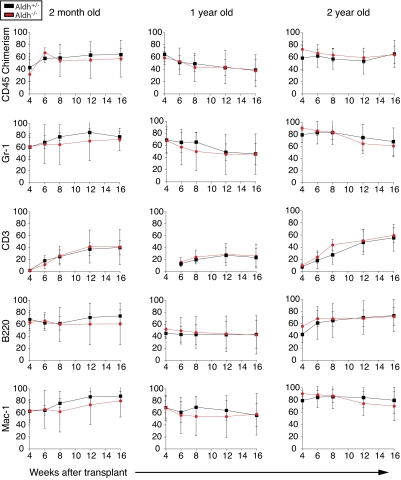
High levels of aldehyde dehydrogenase (ALDH) activity have been proposed to be a common feature of stem cells. Adult hematopoietic, neural, and cancer stem cells have all been reported to have high ALDH activity, detected using Aldefluor, a fluorogenic substrate for ALDH. This activity has been attributed to Aldh1a1, an enzyme that is expressed at high levels in stem cells and that has been suggested to regulate stem cell function. Nonetheless, Aldh1a1 function in stem cells has never been tested genetically. We observed that Aldh1a1 was preferentially expressed in mouse hematopoietic stem cells (HSCs) and expression increased with age. Hematopoietic cells from Aldh1a1-deficient mice exhibited increased sensitivity to cyclophosphamide in a non-cell-autonomous manner, consistent with its role in cyclophosphamide metabolism in the liver. However, Aldh1a1 deficiency did not affect hematopoiesis, HSC function, or the capacity to reconstitute irradiated recipients in young or old adult mice. Aldh1a1 deficiency also did not affect Aldefluor staining of hematopoietic cells. Finally, Aldh1a1 deficiency did not affect the function of stem cells from the adult central or peripheral nervous systems. Aldh1a1 is not a critical regulator of adult stem cell function or Aldefluor staining in mice.
Figures
 ) and Aldh1a1−/− (
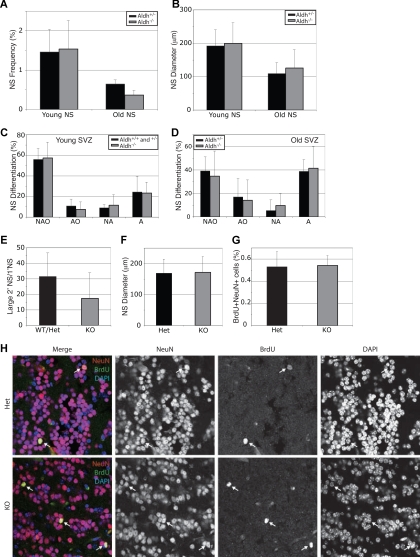
) and Aldh1a1−/− ( ) mice were cultured. Aldh1a1 deficiency did not affect the frequency of neurospheres (A), neurosphere diameter after 8 to 10 days in culture (B), neurosphere differentiation (C,D), or the capacity of primary neurospheres to form large (> 100 μm; these are almost always multipotent) secondary neurospheres upon subcloning (E). Colonies were assessed for the presence of neurons (N), astrocytes (A) and oligodendrocytes (O). The size of secondary neurospheres was also not affected (F). All data represent a total of 3 to 7 mice per treatment, in at least 3 independent experiments except for F which represents 2 independent experiments with 1 mouse per experiment. (G, H) Aldh1a1 deficiency also did not affect the rate of neurogenesis (frequency of BrdU + NeuN + neurons) in the olfactory bulb of 18-month-old adult mice in vivo (n = 2 mice with 25 sections per mouse). Error bars represent SD.
) mice were cultured. Aldh1a1 deficiency did not affect the frequency of neurospheres (A), neurosphere diameter after 8 to 10 days in culture (B), neurosphere differentiation (C,D), or the capacity of primary neurospheres to form large (> 100 μm; these are almost always multipotent) secondary neurospheres upon subcloning (E). Colonies were assessed for the presence of neurons (N), astrocytes (A) and oligodendrocytes (O). The size of secondary neurospheres was also not affected (F). All data represent a total of 3 to 7 mice per treatment, in at least 3 independent experiments except for F which represents 2 independent experiments with 1 mouse per experiment. (G, H) Aldh1a1 deficiency also did not affect the rate of neurogenesis (frequency of BrdU + NeuN + neurons) in the olfactory bulb of 18-month-old adult mice in vivo (n = 2 mice with 25 sections per mouse). Error bars represent SD. ) and Aldh1a1−/− () mice were cultured. Aldh1a1 deficiency did not affect the frequency of neurospheres (A), neurosphere diameter after 9 to 11 days in culture (B), neurosphere differentiation (C,D), or the capacity of primary neurospheres to form multipotent secondary neurospheres upon subcloning (E). Colonies were assessed for the presence of neurons (N), glia (G), and myofibroblasts (M). All data represent a total of 3 to 8 mice per treatment, in at least 3 independent experiments. Error bars represent SD.
) and Aldh1a1−/− () mice were cultured. Aldh1a1 deficiency did not affect the frequency of neurospheres (A), neurosphere diameter after 9 to 11 days in culture (B), neurosphere differentiation (C,D), or the capacity of primary neurospheres to form multipotent secondary neurospheres upon subcloning (E). Colonies were assessed for the presence of neurons (N), glia (G), and myofibroblasts (M). All data represent a total of 3 to 8 mice per treatment, in at least 3 independent experiments. Error bars represent SD.References
-
- Duester G. Genetic dissection of retinoid dehydrogenases. Chem Biol Interact. 2001;130–132:469–480. - PubMed
-
- Niederreither K, Subbarayan V, Dolle P, Chambon P. Embryonic retinoic acid synthesis is essential for early mouse post-implantation development. Nat Genet. 1999;21:444–448. - PubMed
-
- Colvin M, Russo JE, Hilton J, Dulik DM, Fenselau C. Enzymatic mechanisms of resistance to alkylating agents in tumor cells and normal tissues. Adv Enzyme Regul. 1988;27:211–221. - PubMed
-
- Hilton J. Role of aldehyde dehydrogenase in cyclophosphamide-resistant L1210 leukemia. Cancer Res. 1984;44:5156–5160. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
