

CNS learns stable, accurate, and efficient movements using a simple algorithm
- PMID: 18971459
- PMCID: PMC6671516
- DOI: 10.1523/JNEUROSCI.3099-08.2008
CNS learns stable, accurate, and efficient movements using a simple algorithm
Abstract
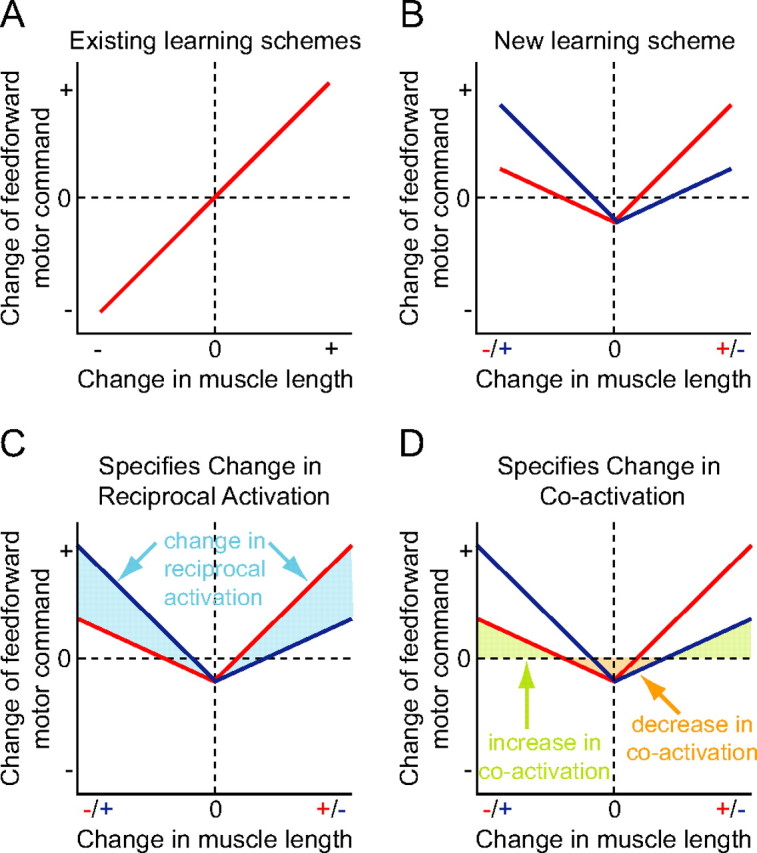
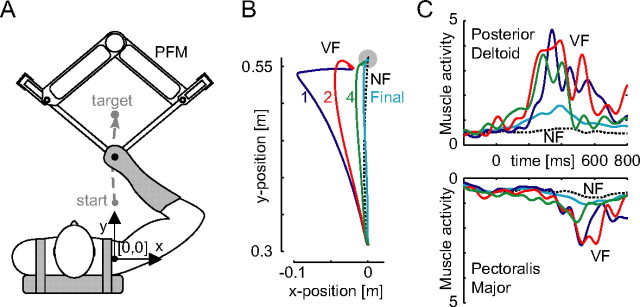
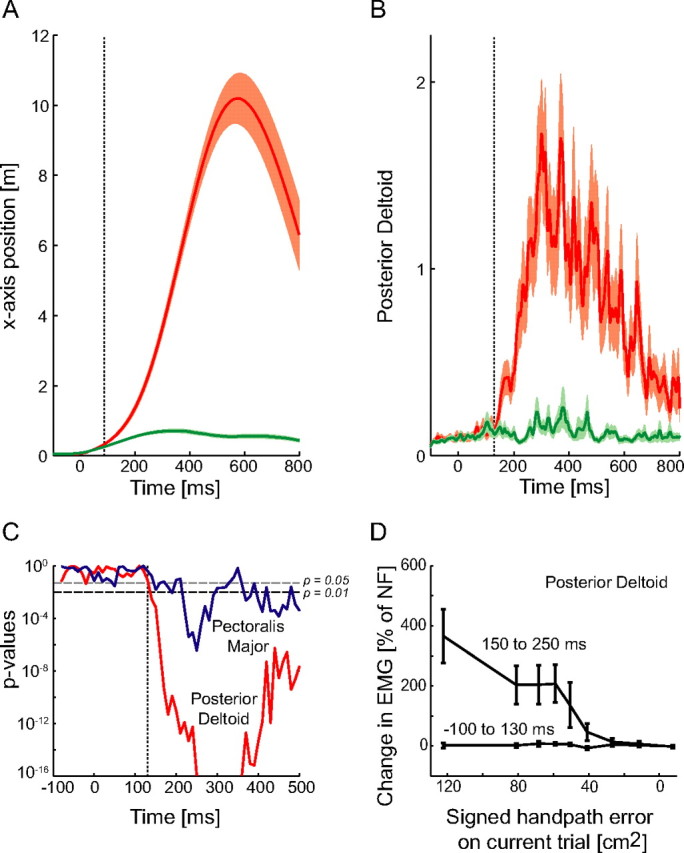
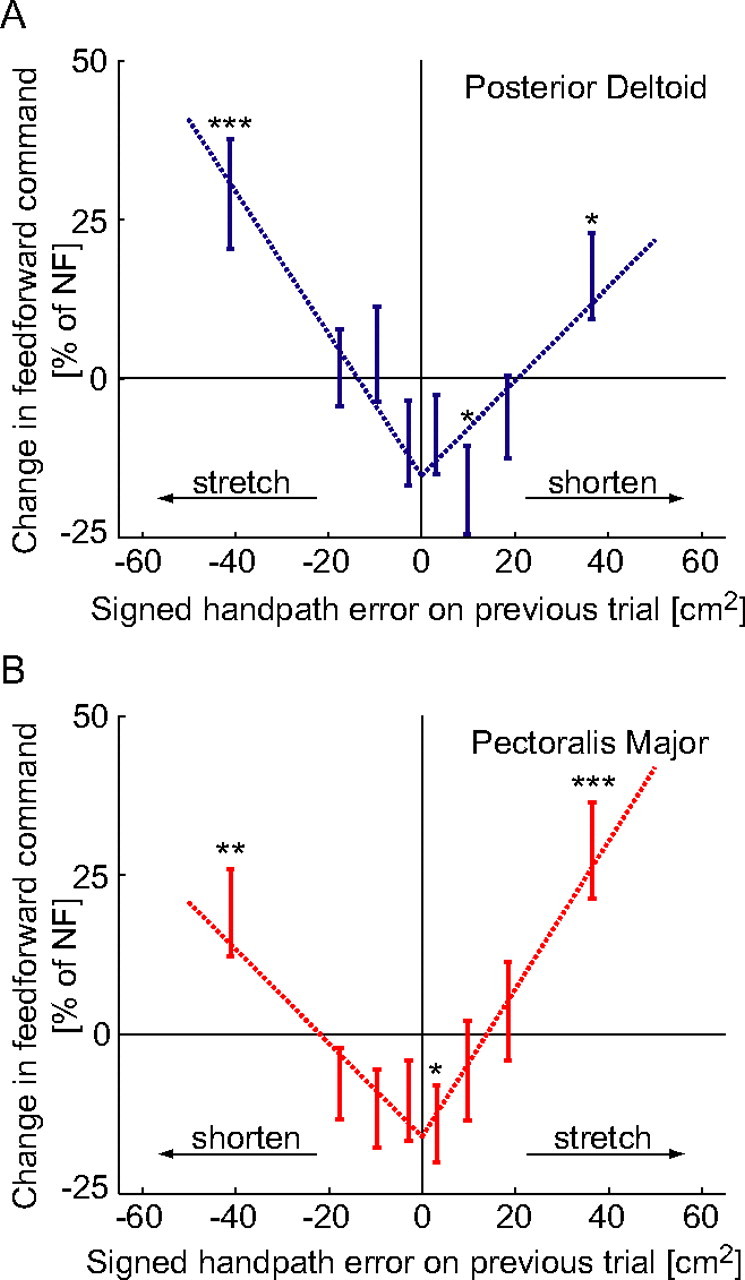
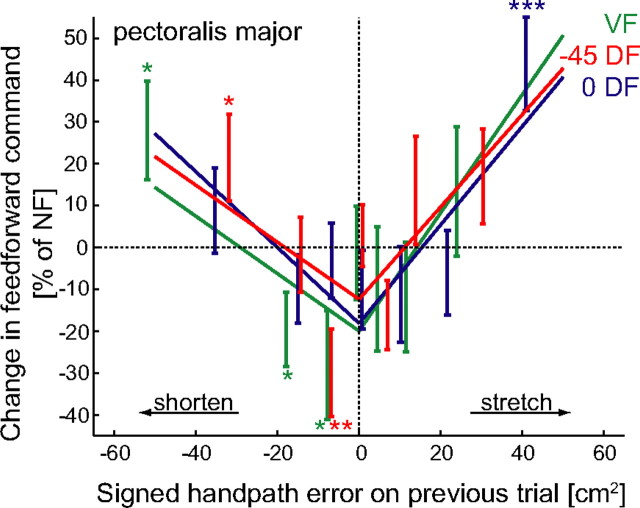
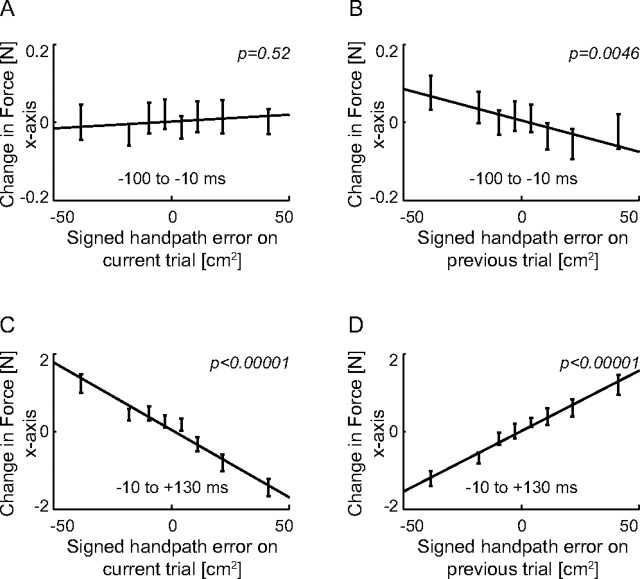
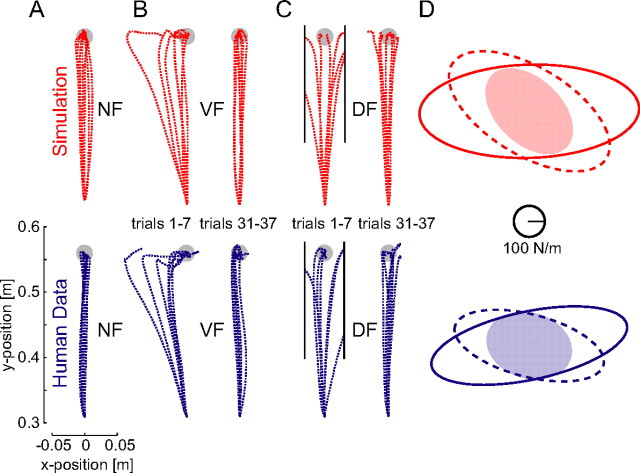
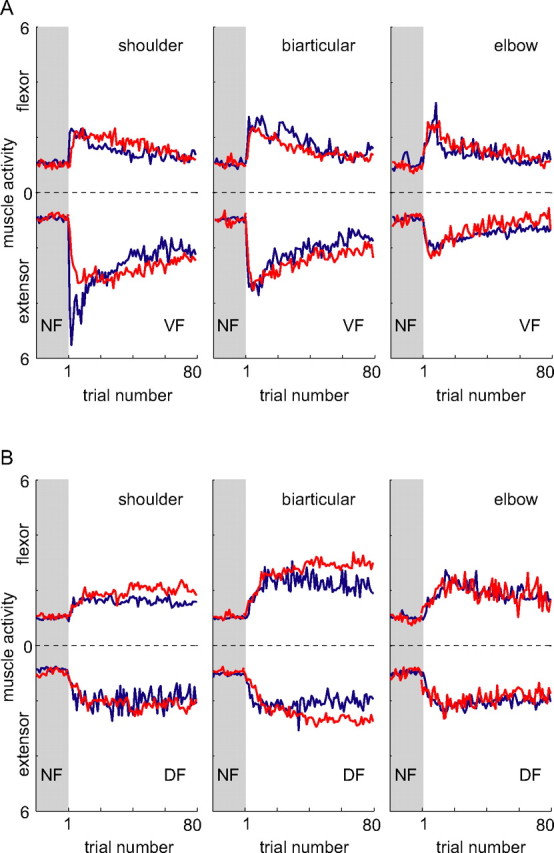
We propose a new model of motor learning to explain the exceptional dexterity and rapid adaptation to change, which characterize human motor control. It is based on the brain simultaneously optimizing stability, accuracy and efficiency. Formulated as a V-shaped learning function, it stipulates precisely how feedforward commands to individual muscles are adjusted based on error. Changes in muscle activation patterns recorded in experiments provide direct support for this control scheme. In simulated motor learning of novel environmental interactions, muscle activation, force and impedance evolved in a manner similar to humans, demonstrating its efficiency and plausibility. This model of motor learning offers new insights as to how the brain controls the complex musculoskeletal system and iteratively adjusts motor commands to improve motor skills with practice.
Figures
References
-
- Burdet E, Milner TE. Quantization of human motions and learning of accurate movements. Biol Cybern. 1998;78:307–318. - PubMed
-
- Burdet E, Codourey A, Rey L. Experimental evaluation of nonlinear adaptive controllers. IEEE Control Syst Mag. 1998;18:39–47.
-
- Burdet E, Osu R, Franklin DW, Yoshioka T, Milner TE, Kawato M. A method for measuring endpoint stiffness during multi-joint arm movements. J Biomech. 2000;33:1705–1709. - PubMed
-
- Burdet E, Osu R, Franklin DW, Milner TE, Kawato M. The central nervous system stabilizes unstable dynamics by learning optimal impedance. Nature. 2001;414:446–449. - PubMed
-
- Burdet E, Tee KP, Mareels I, Milner TE, Chew CM, Franklin DW, Osu R, Kawato M. Stability and motor adaptation in human arm movements. Biol Cybern. 2006;94:20–32. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources