

Brain metabolism dictates the polarity of astrocyte control over arterioles
- PMID: 18971930
- PMCID: PMC4097022
- DOI: 10.1038/nature07525
Brain metabolism dictates the polarity of astrocyte control over arterioles
Abstract
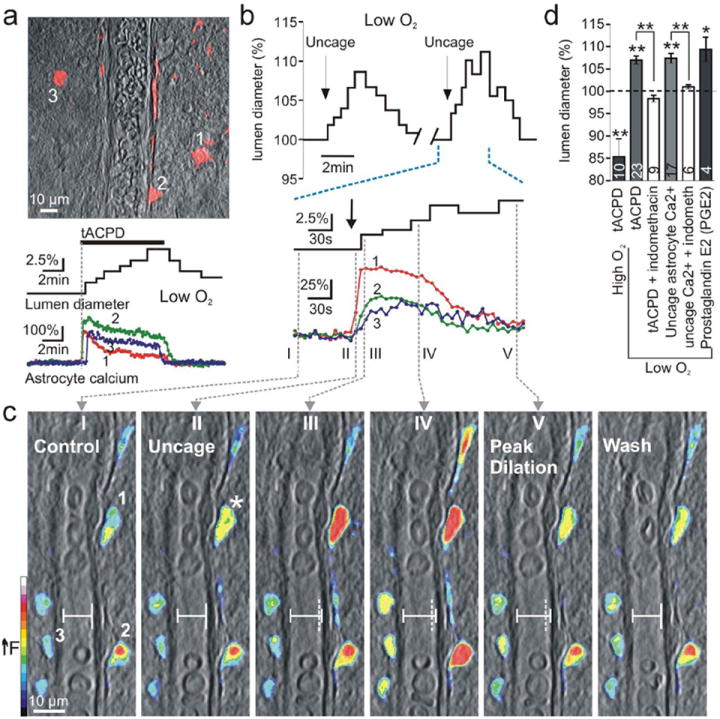
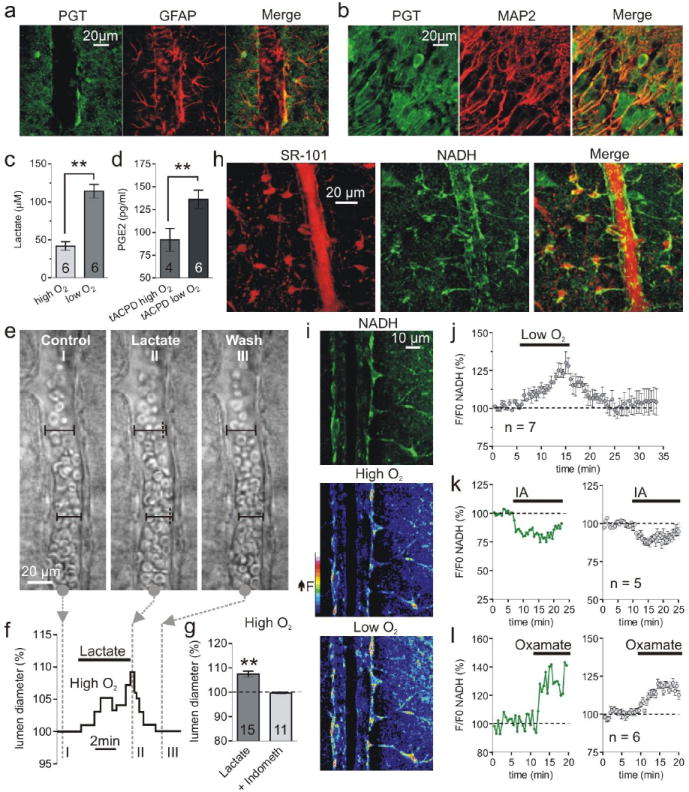
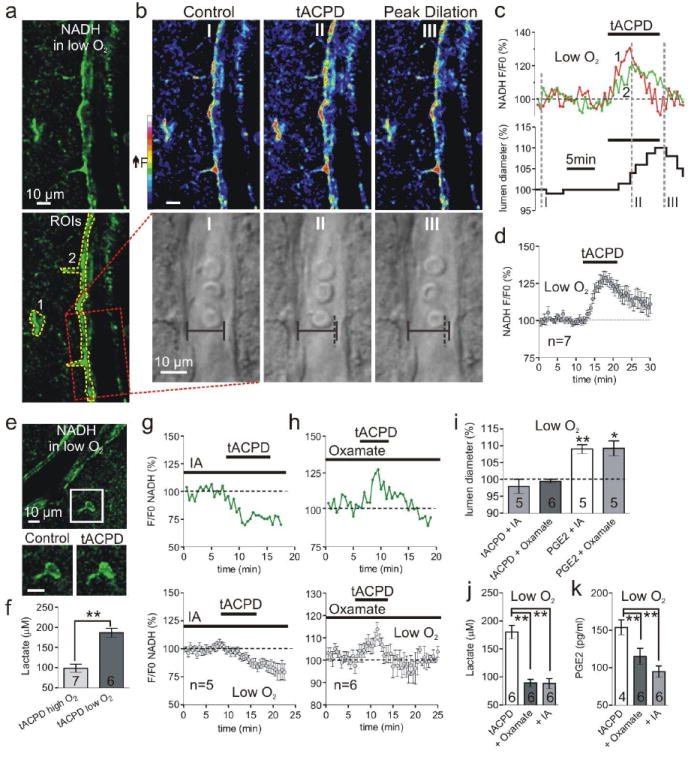
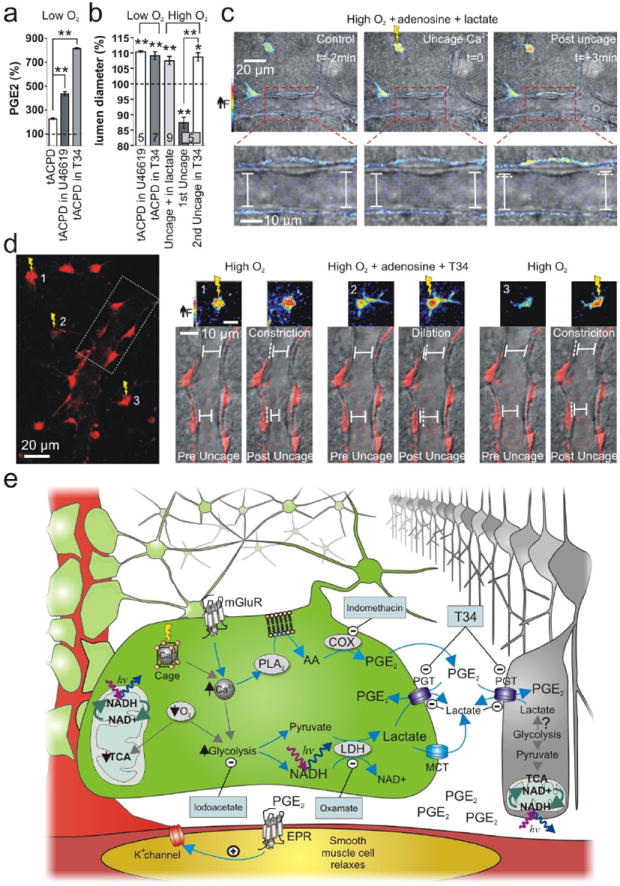
Calcium signalling in astrocytes couples changes in neural activity to alterations in cerebral blood flow by eliciting vasoconstriction or vasodilation of arterioles. However, the mechanism for how these opposite astrocyte influences provide appropriate changes in vessel tone within an environment that has dynamic metabolic requirements remains unclear. Here we show that the ability of astrocytes to induce vasodilations over vasoconstrictions relies on the metabolic state of the rat brain tissue. When oxygen availability is lowered and astrocyte calcium concentration is elevated, astrocyte glycolysis and lactate release are maximized. External lactate attenuates transporter-mediated uptake from the extracellular space of prostaglandin E(2), leading to accumulation and subsequent vasodilation. In conditions of low oxygen concentration extracellular adenosine also increases, which blocks astrocyte-mediated constriction, facilitating dilation. These data reveal the role of metabolic substrates in regulating brain blood flow and provide a mechanism for differential astrocyte control over cerebrovascular diameter during different states of brain activation.
Figures
Comment in
-
Neuroscience: Brain power.Nature. 2008 Dec 11;456(7223):715-6. doi: 10.1038/456715a. Nature. 2008. PMID: 19079043 No abstract available.
References
-
- Mulligan SJ, MacVicar BA. Calcium transients in astrocyte endfeet cause cerebrovascular constrictions. Nature. 2004;431(7005):195. - PubMed
-
- Zonta M, Angulo MC, Gobbo S, et al. Neuron-to-astrocyte signaling is central to the dynamic control of brain microcirculation. Nature neuroscience. 2003;6(1):43. - PubMed
-
- Filosa JA, Bonev AD, Nelson MT. Calcium dynamics in cortical astrocytes and arterioles during neurovascular coupling. Circulation research. 2004;95(10):e73. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
