

Conservation and diversity of Foxp2 expression in muroid rodents: functional implications
- PMID: 18972576
- PMCID: PMC2677918
- DOI: 10.1002/cne.21881
Conservation and diversity of Foxp2 expression in muroid rodents: functional implications
Abstract
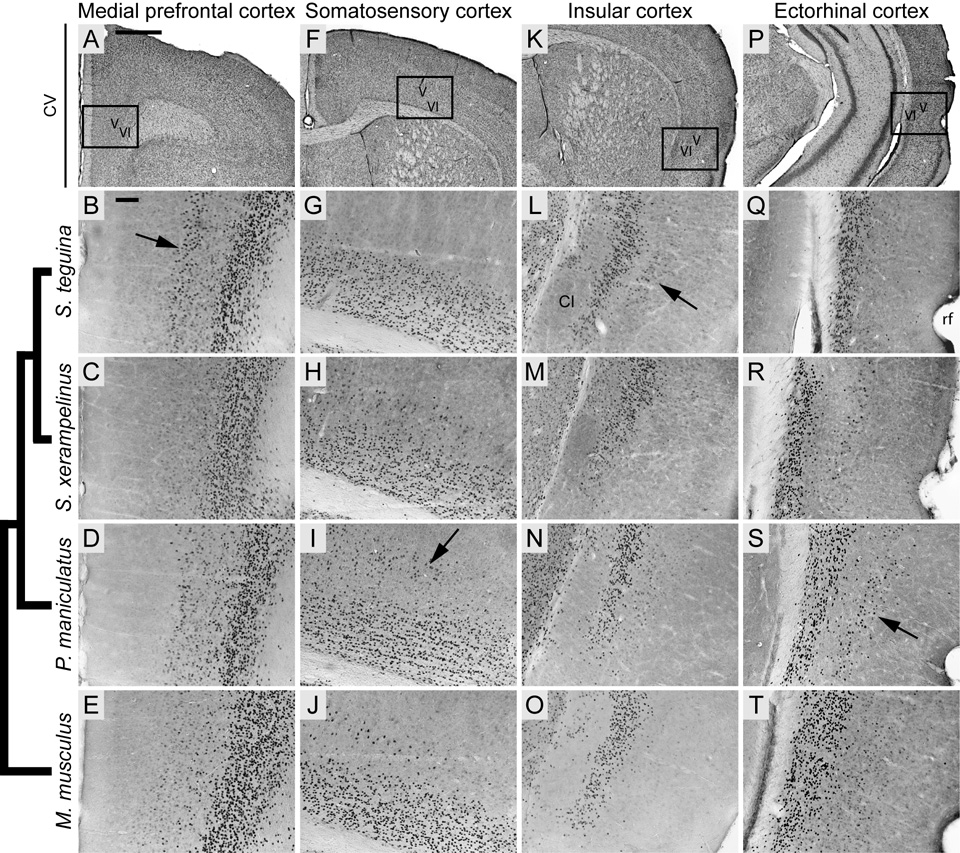
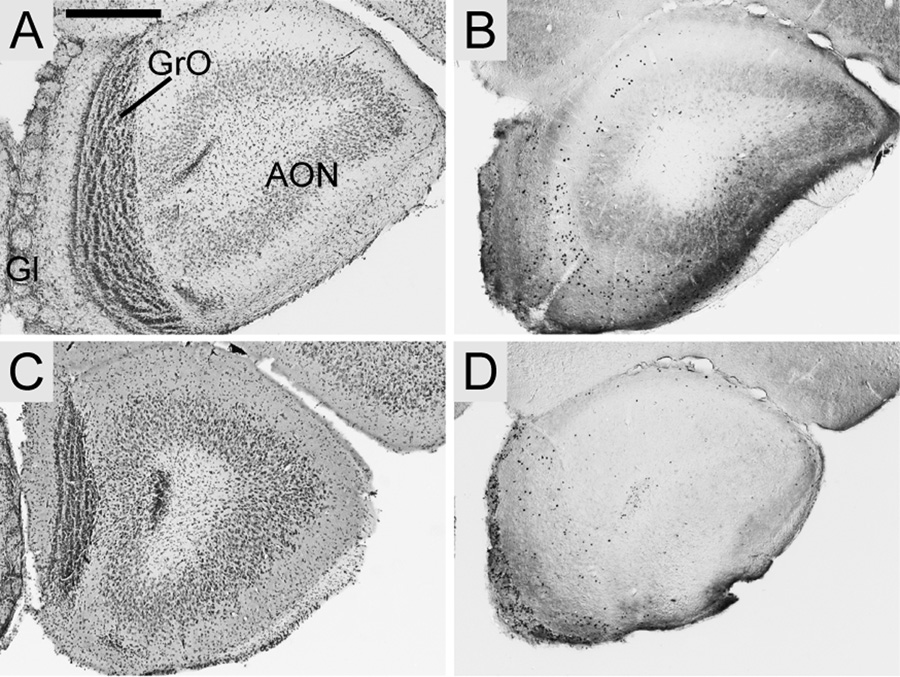
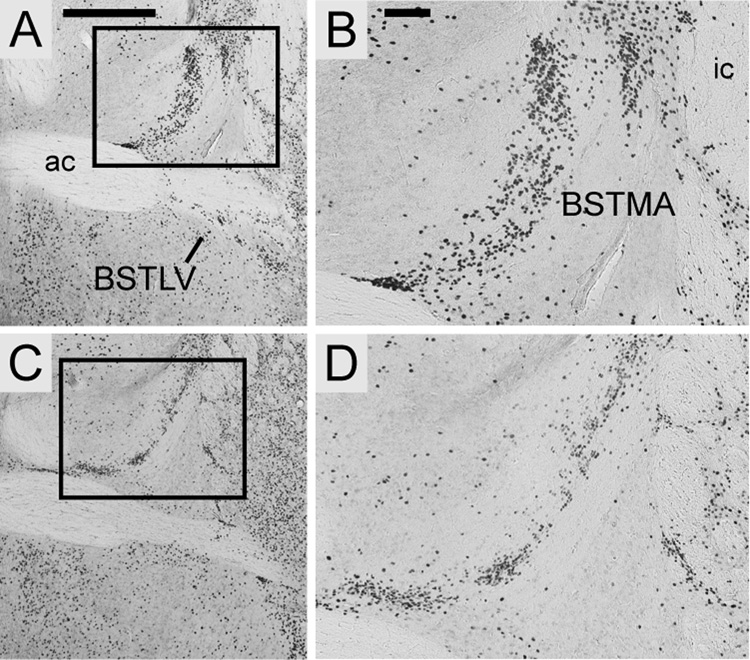
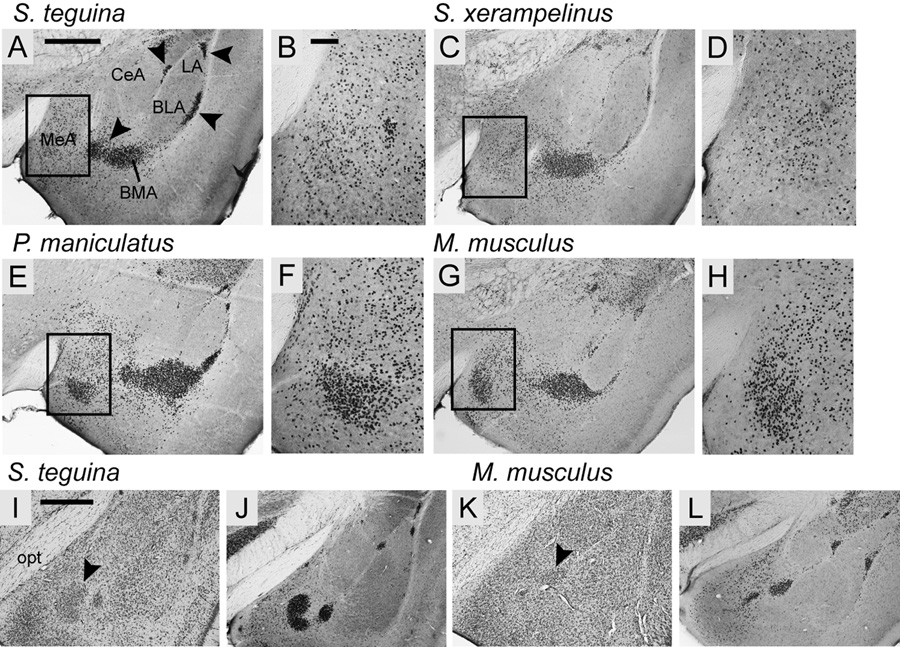
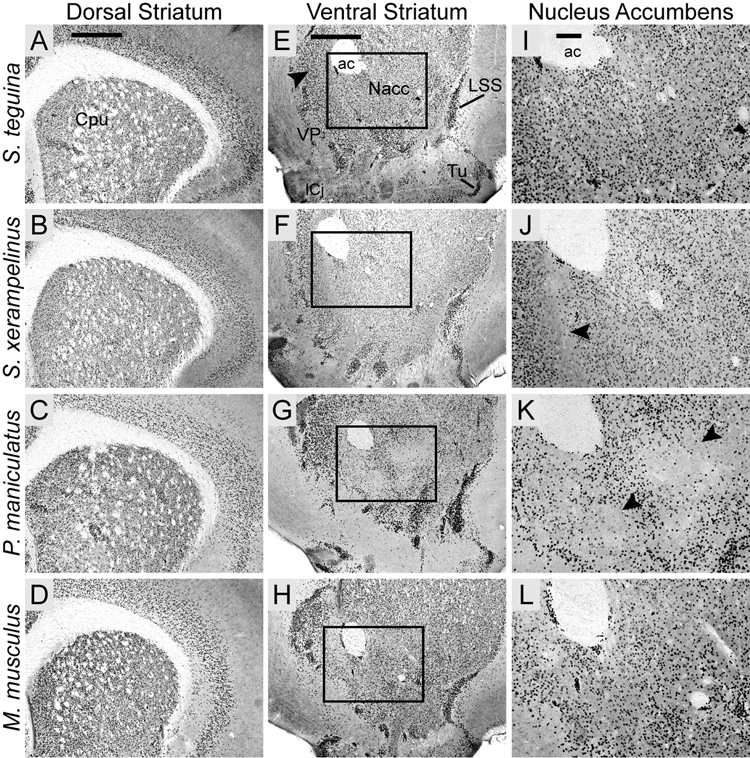
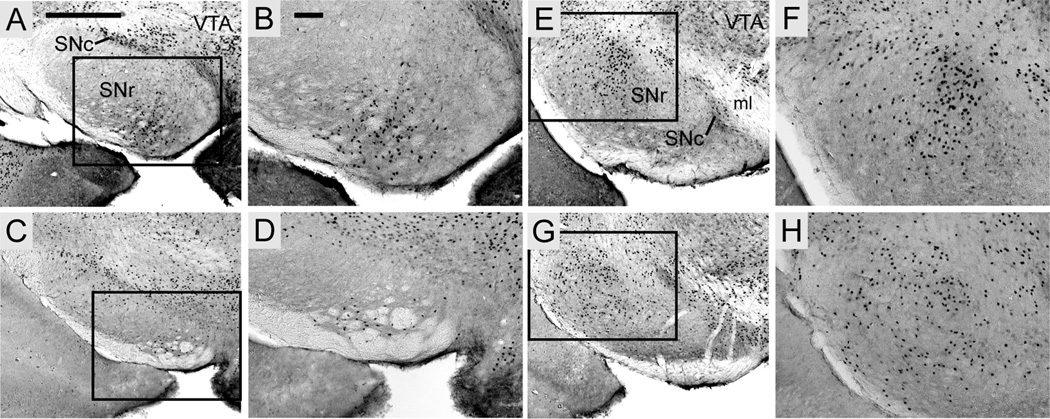
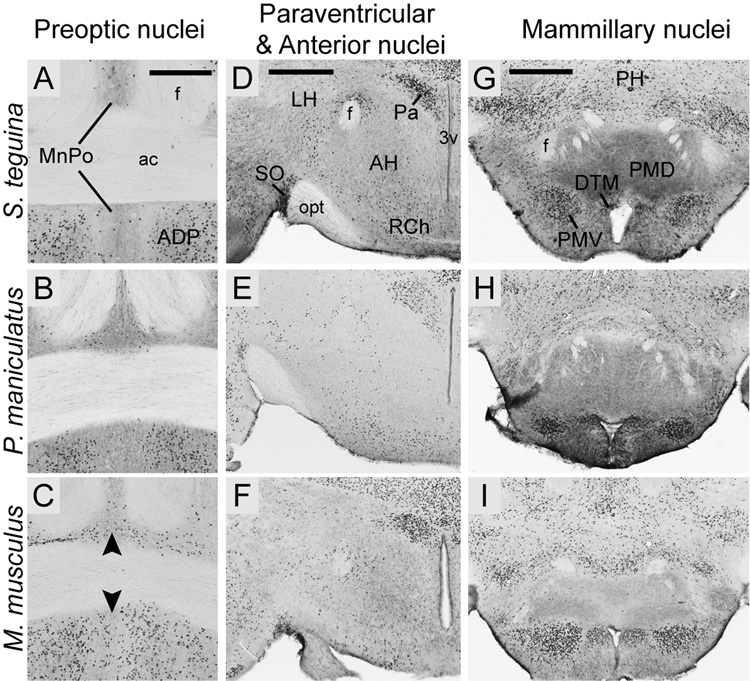
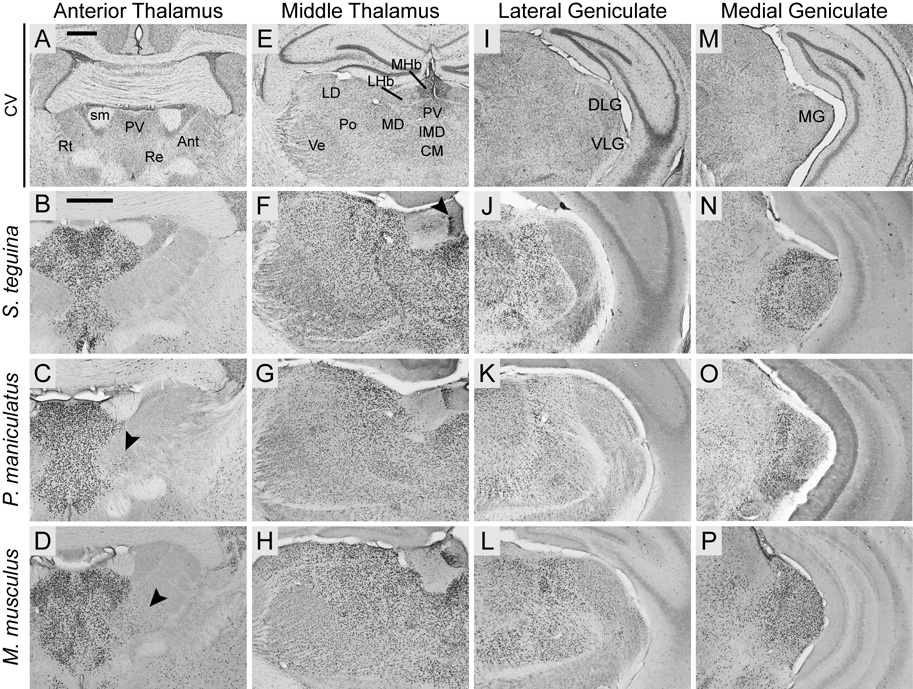
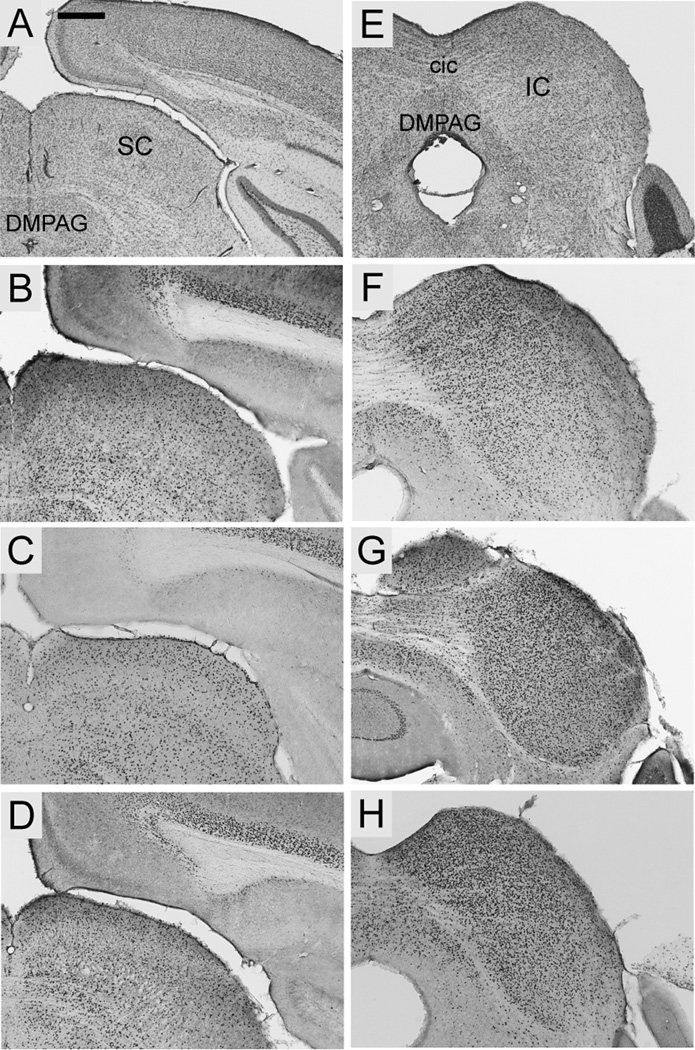
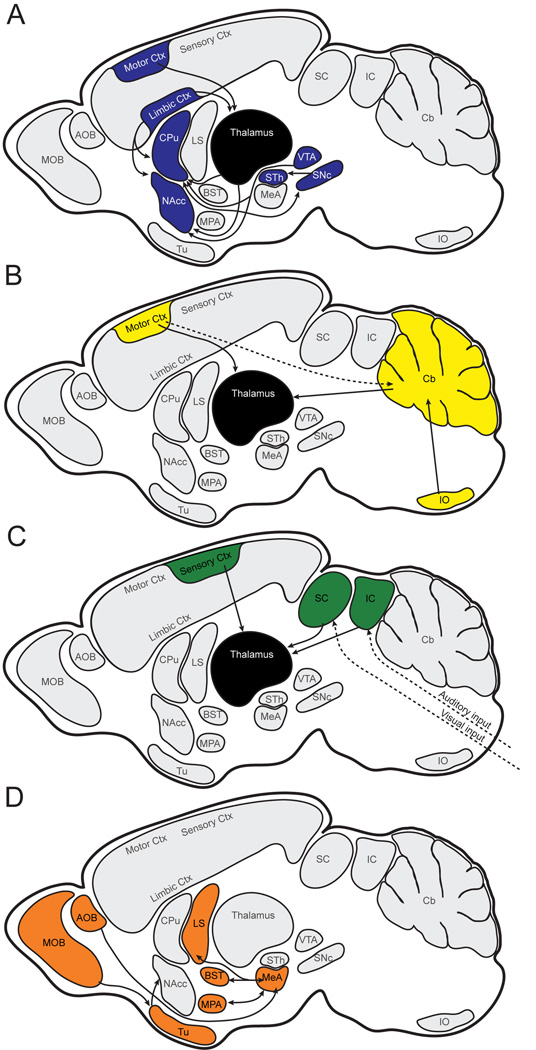
FOXP2, the first gene causally linked to a human language disorder, is implicated in song acquisition, production, and perception in oscine songbirds, the evolution of speech and language in hominids, and the evolution of echolocation in bats. Despite the evident relevance of Foxp2 to vertebrate acoustic communication, a comprehensive description of neural expression patterns is currently lacking in mammals. Here we use immunocytochemistry to systematically describe the neural distribution of Foxp2 protein in four species of muroid rodents: Scotinomys teguina and S. xerampelinus ("singing mice"), the deer mouse, Peromyscus maniculatus, and the lab mouse, Mus musculus. While expression patterns were generally highly conserved across brain regions, we identified subtle but consistent interspecific differences in Foxp2 distribution, most notably in the medial amygdala and nucleus accumbens, and in layer V cortex throughout the brain. Throughout the brain, Foxp2 was highly enriched in areas involved in modulation of fine motor output (striatum, mesolimbic dopamine circuit, olivocerebellar system) and in multimodal sensory processing and sensorimotor integration (thalamus, cortex). We propose a generalized model for Foxp2-modulated pathways in the adult brain including, but not limited to, fine motor production and auditory perception.
Figures


References
-
- Ackermann H, Graber S, Hertrich I, Daum I. Cerebellar contributions to the perception of temporal cues within the speech and nonspeech domain. Brain and Language. 1999;67:228–241. - PubMed
-
- Alcock KJ, Passingham RE, Watkins K, Vargha-Khadem F. Pitch and timing abilities in inherited speech and language impairment. Brain and Language. 2000;75:34–46. - PubMed
-
- Alloway KD, Lou L, Nwabueze-Ogbo F, Chakrabarti S. Topography of cortical projections to the dorsolateral neostriatum in rats: multiple overlapping sensorimotor pathways. Journal of Comparative Neurology. 2006;499:33–48. - PubMed
-
- Bayer SA. Neurogenetic patterns in the medial limbic cortex of the rat related to anatomical connections with the thalamus and striatum. Experimental Neurology. 1990;107:132–142. - PubMed
-
- Behbehani MM. Functional characteristics of the midbrain periaqueductal gray. Progress in Neurobiology. 1995;46:575–605. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
