

Cholesterol feedback: from Schoenheimer's bottle to Scap's MELADL
- PMID: 18974038
- PMCID: PMC2674699
- DOI: 10.1194/jlr.R800054-JLR200
Cholesterol feedback: from Schoenheimer's bottle to Scap's MELADL
Abstract
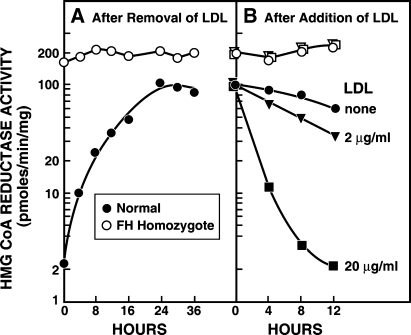
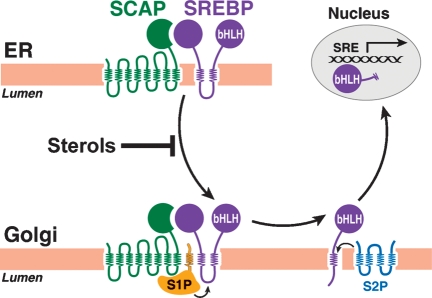
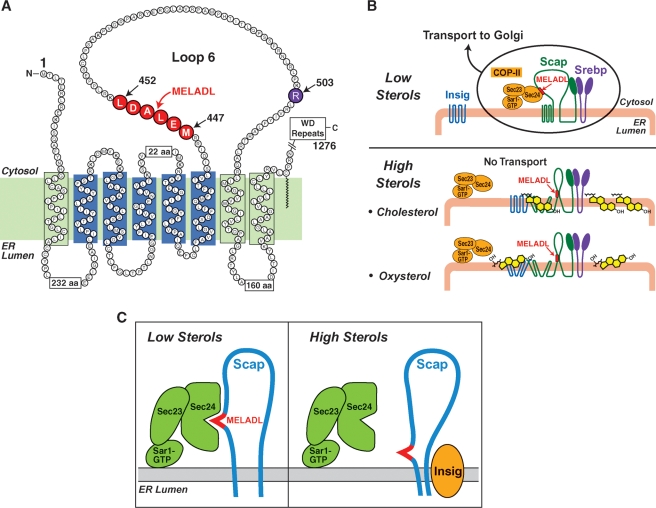
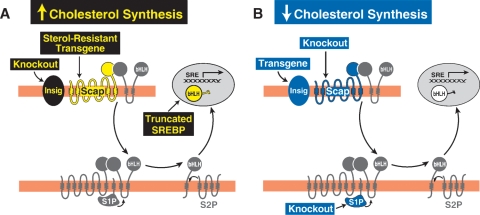
Cholesterol biosynthesis is among the most intensely regulated processes in biology. Synthetic rates vary over hundreds of fold depending on the availability of an external source of cholesterol. Studies of this feedback regulatory process have a rich history. The field began 75 years ago when Rudolf Schoenheimer measured cholesterol balance in mice in a bottle. He found that cholesterol feeding led to decreased cholesterol synthesis, thereby introducing the general phenomenon by which end products of biosynthetic pathways inhibit their own synthesis. Recently, cholesterol feedback has been explained at a molecular level with the discovery of membrane-bound transcription factors called sterol regulatory element-binding proteins (SREBPs), and an appreciation of the sterol-sensing role of their partner, an escort protein called Scap. The key element in Scap is a hexapeptide sequence designated MELADL (rhymes with bottle). Thus, over 75 years, Schoenheimer's bottle led to Scap's MELADL. In addition to their basic importance in membrane biology, these studies have implications for the regulation of plasma cholesterol levels and consequently for the development of atherosclerotic plaques, myocardial infarctions, and strokes. In this article we review the major milestones in the cholesterol feedback story.
Figures

References
-
- Schoenheimer R., and F. Breusch. 1933. Synthesis and destruction of cholesterol in the organism. J. Biol. Chem. 103 439–448.
-
- Jacob F., and J. Monod. 1961. On the regulation of gene activity. Cold Spring Harb. Symp. Quant. Biol. 26 193–211. - PubMed
-
- Bloch K. 1965. The biological synthesis of cholesterol. Science. 50 19–128. - PubMed
-
- Gould R. G., C. B. Taylor, J. S. Hagerman, I. Warner, and D. J. Campbell. 1953. Cholesterol metabolism: I. Effect of dietary cholesterol on the synthesis of cholesterol in dog tissue in vitro. J. Biol. Chem. 201 519–523. - PubMed
-
- Spady D. K., and J. M. Dietschy. 1983. Sterol synthesis in vivo in 18 tissues of the squirrel monkey, guinea pig, rabbit, hamster, and rat. J. Lipid Res. 24 303–315. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
