

Aurora kinase A inhibition leads to p73-dependent apoptosis in p53-deficient cancer cells
- PMID: 18974145
- PMCID: PMC2587495
- DOI: 10.1158/0008-5472.CAN-08-2658
Aurora kinase A inhibition leads to p73-dependent apoptosis in p53-deficient cancer cells
Abstract
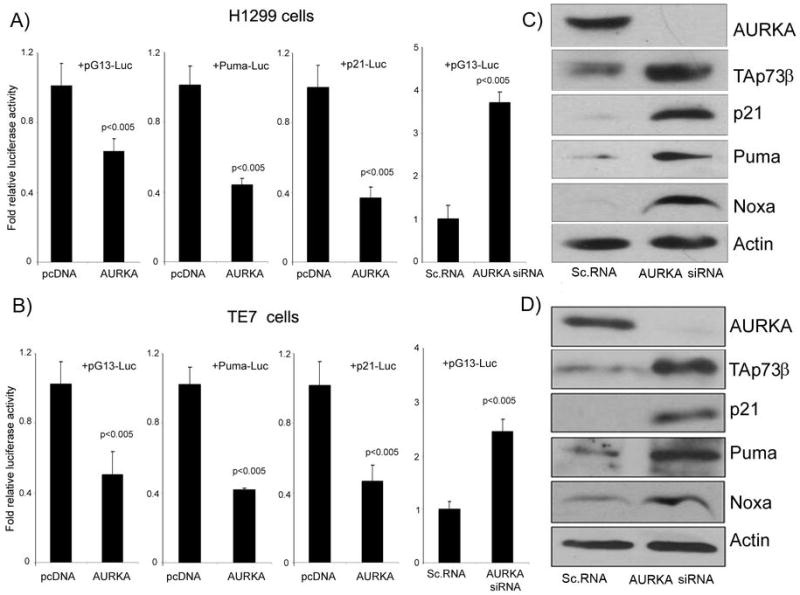
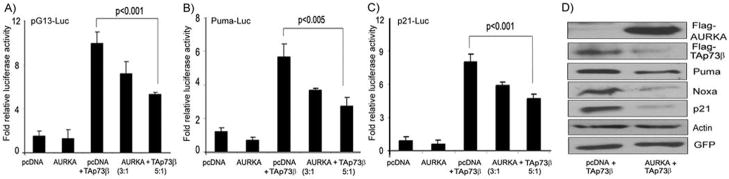
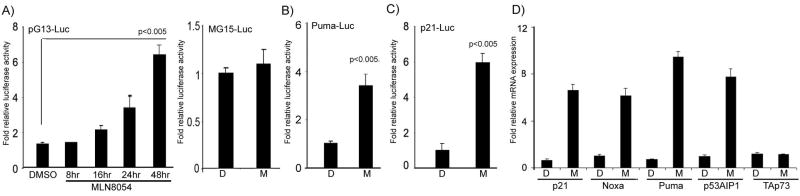
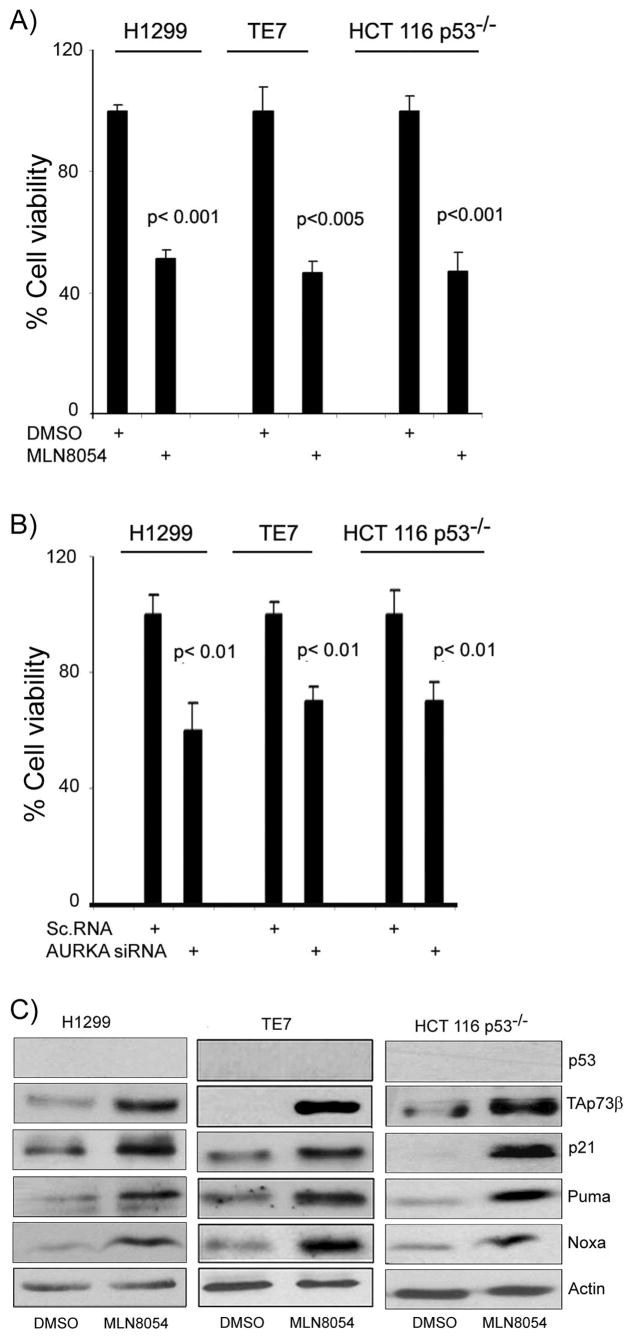
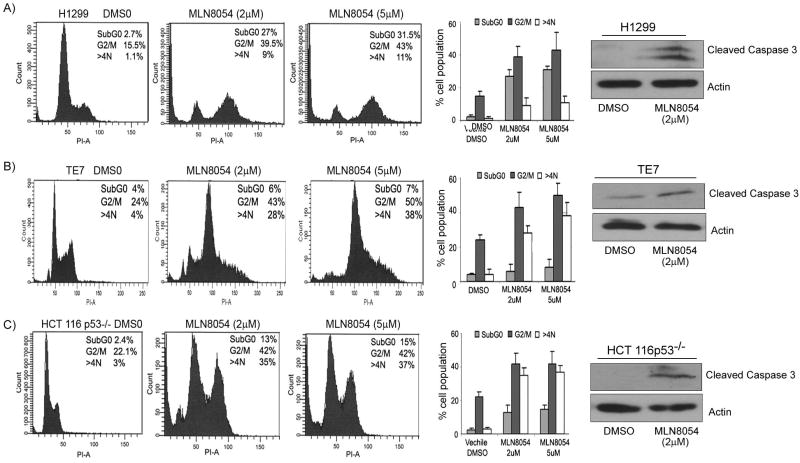
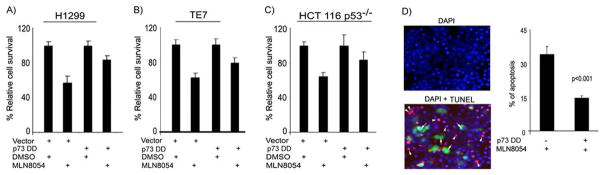
We investigated the role of Aurora kinase A (AURKA) in regulating p73-dependent apoptosis using the p53-deficient cancer cell lines H1299, TE7, and HCT116p53(-/-). Overexpression of AURKA led to down-regulation of the TAp73-induced activation of the p53/p73-dependent luciferase reporter plasmid (pG13-luc). The reduction in the TAp73 transcription activity was confirmed by measuring the activity of luciferase reporters for p21/WAF1, and PUMA. The siRNA knockdown of endogenous AURKA reversed these effects and Western blot analysis showed a significant increase in the protein level of TAp73 and its downstream transcription targets, PUMA, NOXA, and p21/WAF1. The coexpression of AURKA together with TAp73 inhibited the activation of the pG13-luc, PUMA-luc, and p21/WAF1-luc reporter plasmids with reduction in the protein levels of TAp73 transcription targets. Treatment with AURKA-selective small molecule inhibitor MLN8054 led to a significant increase in the activities of pG13-luc, PUMA-luc, and p21/WAF1-luc reporter plasmids. This effect was accompanied by a significant increase in the mRNA and protein levels of several TAp73 transcription targets: p21/WAF1, PUMA, and NOXA. Flow cytometry cell cycle analysis, after MLN8054 treatment, showed more than a 2-fold increase in cell death. The apoptotic outcome was corroborated by showing an increase in cleaved caspase-3 protein levels by Western blot. Using terminal deoxynucleotidyl-transferase-mediated dUTP nick-end labeling assay, we showed that the expression of dominant-negative mutant TAp73 expression plasmid (p73DD) counteracted the MLN8054-induced cell death. Taken together, our results indicate that AURKA regulates TAp73-dependent apoptosis and highlight the potential of the AURKA inhibitor MLN8054 in treating cancers that are defective in p53 signaling.
Figures
References
-
- Zhou H, Kuang J, Zhong L, et al. Tumour amplified kinase STK15/BTAK induces centrosome amplification, aneuploidy and transformation. Nat Genet. 1998;20:189–93. - PubMed
-
- Glover DM, Leibowitz MH, McLean DA, Parry H. Mutations in aurora prevent centrosome separation leading to the formation of monopolar spindles. Cell. 1995;81:95–105. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
