

Genome 3'-end repair in dengue virus type 2
- PMID: 18974278
- PMCID: PMC2590968
- DOI: 10.1261/rna.1051208
Genome 3'-end repair in dengue virus type 2
Abstract
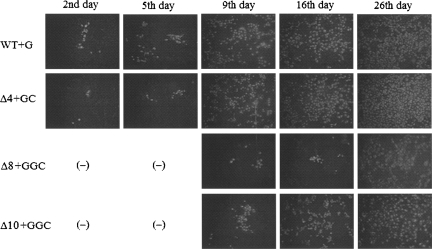
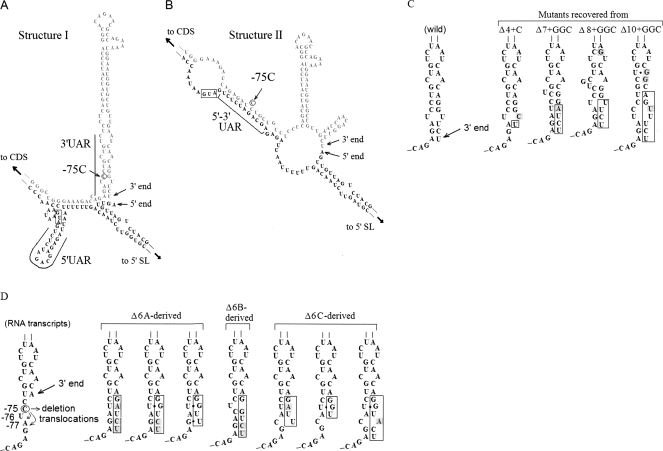
Genomes of RNA viruses encounter a continual threat from host cellular ribonucleases. Therefore, viruses have evolved mechanisms to protect the integrity of their genomes. To study the mechanism of 3'-end repair in dengue virus-2 in mammalian cells, a series of 3'-end deletions in the genome were evaluated for virus replication by detection of viral antigen NS1 and by sequence analysis. Limited deletions did not cause any delay in the detection of NS1 within 5 d. However, deletions of 7-10 nucleotides caused a delay of 9 d in the detection of NS1. Sequence analysis of RNAs from recovered viruses showed that at early times, virus progenies evolved through RNA molecules of heterogeneous lengths and nucleotide sequences at the 3' end, suggesting a possible role for terminal nucleotidyl transferase activity of the viral polymerase (NS5). However, this diversity gradually diminished and consensus sequences emerged. Template activities of 3'-end mutants in the synthesis of negative-strand RNA in vitro by purified NS5 correlate well with the abilities of mutant RNAs to repair and produce virus progenies. Using the Mfold program for RNA structure prediction, we show that if the 3' stem-loop (3' SL) structure was abrogated by mutations, viruses eventually restored the 3' SL structure. Taken together, these results favor a two-step repair process: non-template-based nucleotide addition followed by evolutionary selection of 3'-end sequences based on the best-fit RNA structure that can support viral replication.
Figures


Similar articles
-
Identification of specific nucleotide sequences within the conserved 3'-SL in the dengue type 2 virus genome required for replication.J Virol. 1998 Sep;72(9):7510-22. doi: 10.1128/JVI.72.9.7510-7522.1998. J Virol. 1998. PMID: 9696848 Free PMC article.
-
Substitution of NS5 N-terminal domain of dengue virus type 2 RNA with type 4 domain caused impaired replication and emergence of adaptive mutants with enhanced fitness.J Biol Chem. 2014 Aug 8;289(32):22385-400. doi: 10.1074/jbc.M114.584466. Epub 2014 Jun 5. J Biol Chem. 2014. PMID: 24904061 Free PMC article.
-
The 5'-terminal region of the Aichi virus genome encodes cis-acting replication elements required for positive- and negative-strand RNA synthesis.J Virol. 2005 Jun;79(11):6918-31. doi: 10.1128/JVI.79.11.6918-6931.2005. J Virol. 2005. PMID: 15890931 Free PMC article.
-
Functional RNA elements in the dengue virus genome.Viruses. 2011 Sep;3(9):1739-56. doi: 10.3390/v3091739. Epub 2011 Sep 15. Viruses. 2011. PMID: 21994804 Free PMC article. Review.
-
Structural and functional analysis of dengue virus RNA.Novartis Found Symp. 2006;277:120-32; discussion 132-5, 251-3. Novartis Found Symp. 2006. PMID: 17319158 Review.
Cited by
-
Amodiaquine, an antimalarial drug, inhibits dengue virus type 2 replication and infectivity.Antiviral Res. 2014 Jun;106:125-34. doi: 10.1016/j.antiviral.2014.03.014. Epub 2014 Mar 27. Antiviral Res. 2014. PMID: 24680954 Free PMC article.
-
The Golgi associated ERI3 is a Flavivirus host factor.Sci Rep. 2016 Sep 29;6:34379. doi: 10.1038/srep34379. Sci Rep. 2016. PMID: 27682269 Free PMC article.
-
The Pseudo-Circular Genomes of Flaviviruses: Structures, Mechanisms, and Functions of Circularization.Cells. 2021 Mar 13;10(3):642. doi: 10.3390/cells10030642. Cells. 2021. PMID: 33805761 Free PMC article. Review.
-
Molecular basis for nucleotide conservation at the ends of the dengue virus genome.PLoS Pathog. 2012 Sep;8(9):e1002912. doi: 10.1371/journal.ppat.1002912. Epub 2012 Sep 13. PLoS Pathog. 2012. PMID: 23028313 Free PMC article.
-
Infection of Aedes albopictus Mosquito C6/36 Cells with the wMelpop Strain of Wolbachia Modulates Dengue Virus-Induced Host Cellular Transcripts and Induces Critical Sequence Alterations in the Dengue Viral Genome.J Virol. 2019 Jul 17;93(15):e00581-19. doi: 10.1128/JVI.00581-19. Print 2019 Aug 1. J Virol. 2019. PMID: 31092581 Free PMC article.
References
-
- Ackermann M., Padmanabhan R. De novo synthesis of RNA by the dengue virus RNA-dependent RNA polymerase exhibits temperature dependence at the initiation but not elongation phase. J. Biol. Chem. 2001;276:39926–39937. - PubMed
-
- Andrews N.C., Levin D., Baltimore D. Poliovirus replicase stimulation by terminal uridylyl transferase. J. Biol. Chem. 1985;260:7628–7635. - PubMed
-
- Arnold J.J., Ghosh S.K., Cameron C.E. Poliovirus RNA-dependent RNA polymerase (3Dpol): Divalent cation modulation of primer, template, and nucleotide selection. J. Biol. Chem. 1999;274:37060–37069. - PubMed
-
- Brinton M.A., Fernandez A.V., Dispoto J.H. The 3′-nucleotides of flavivirus genomic RNA form a conserved secondary structure. Virology. 1986;153:113–121. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials