

Structural insights into a circadian oscillator
- PMID: 18974343
- PMCID: PMC2588432
- DOI: 10.1126/science.1150451
Structural insights into a circadian oscillator
Abstract
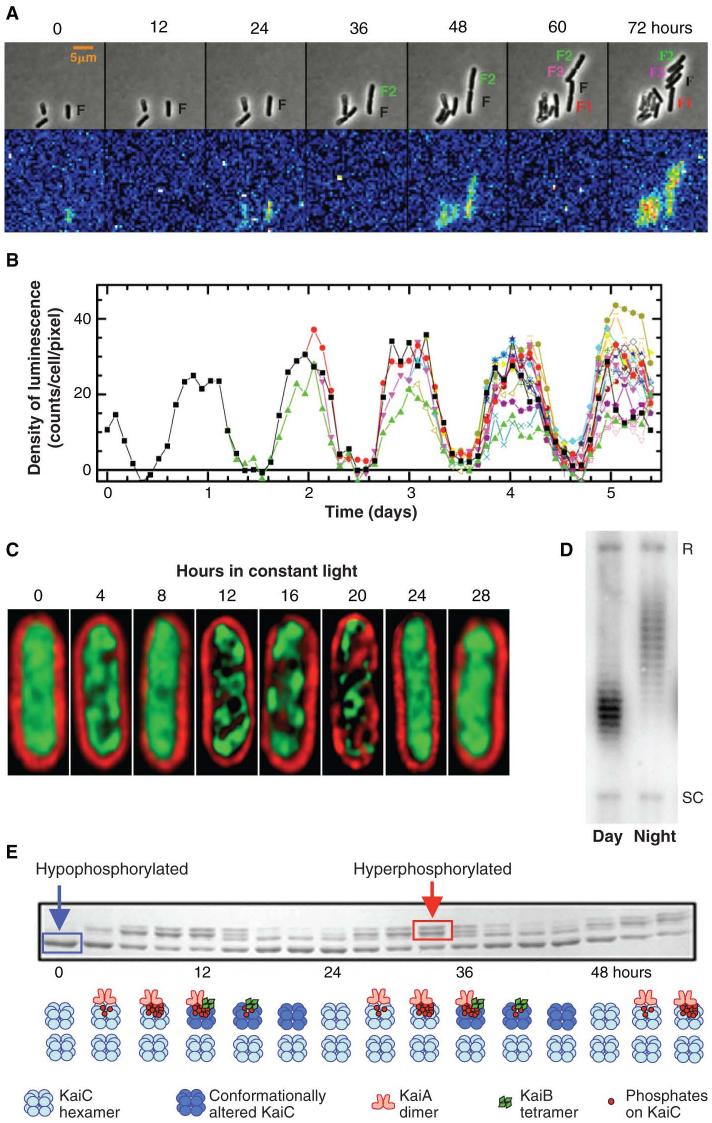
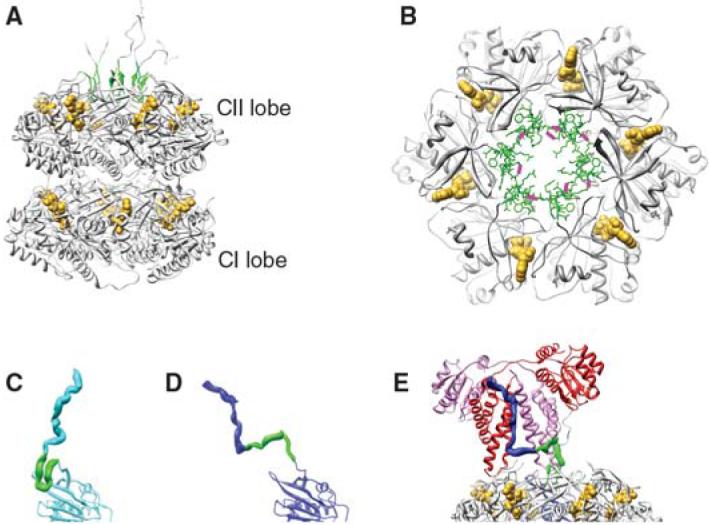
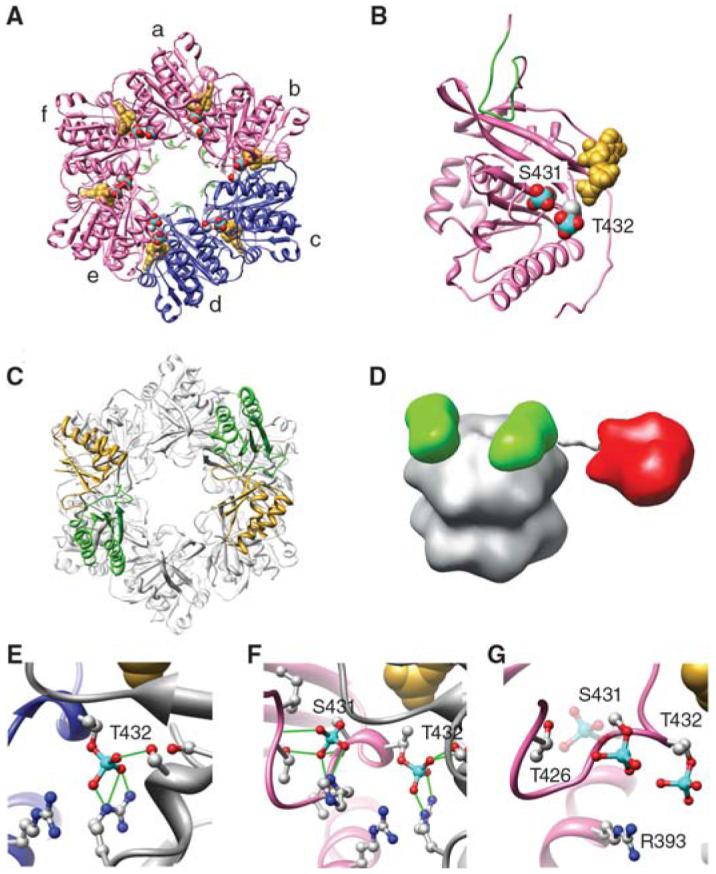
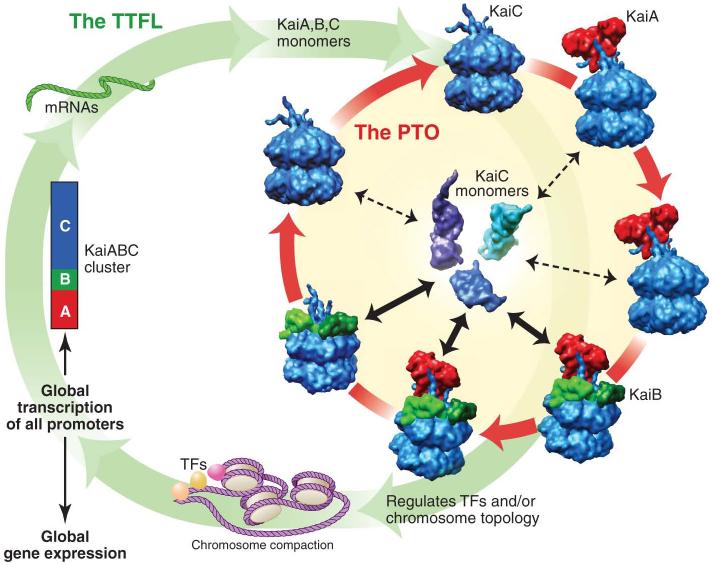
An endogenous circadian system in cyanobacteria exerts pervasive control over cellular processes, including global gene expression. Indeed, the entire chromosome undergoes daily cycles of topological changes and compaction. The biochemical machinery underlying a circadian oscillator can be reconstituted in vitro with just three cyanobacterial proteins, KaiA, KaiB, and KaiC. These proteins interact to promote conformational changes and phosphorylation events that determine the phase of the in vitro oscillation. The high-resolution structures of these proteins suggest a ratcheting mechanism by which the KaiABC oscillator ticks unidirectionally. This posttranslational oscillator may interact with transcriptional and translational feedback loops to generate the emergent circadian behavior in vivo. The conjunction of structural, biophysical, and biochemical approaches to this system reveals molecular mechanisms of biological timekeeping.
Figures
References
-
- Dunlap JC, Loros JJ, DeCoursey PJ, editors. Chronobiology: Biological Timekeeping. Sinauer; Sunderland, MA: 2004.
-
- Mackey SR, Golden SS. Trends Microbiol. 2007;15:381–388. - PubMed
-
- Johnson CH. In: Methods in Enzymology. Young MW, editor. vol. 393. Academic Press; 2005. pp. 818–837. - PubMed
-
- Mihalcescu I, Hsing W, Leibler S. Nature. 2004;430:81–85. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
