

Alteration of the cortical actin cytoskeleton deregulates Ca2+ signaling, monospermic fertilization, and sperm entry
- PMID: 18974786
- PMCID: PMC2570615
- DOI: 10.1371/journal.pone.0003588
Alteration of the cortical actin cytoskeleton deregulates Ca2+ signaling, monospermic fertilization, and sperm entry
Abstract
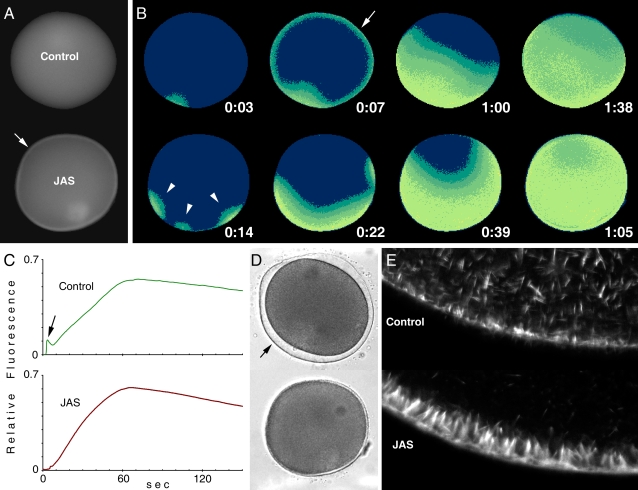
Background: When preparing for fertilization, oocytes undergo meiotic maturation during which structural changes occur in the endoplasmic reticulum (ER) that lead to a more efficient calcium response. During meiotic maturation and subsequent fertilization, the actin cytoskeleton also undergoes dramatic restructuring. We have recently observed that rearrangements of the actin cytoskeleton induced by actin-depolymerizing agents, or by actin-binding proteins, strongly modulate intracellular calcium (Ca2+) signals during the maturation process. However, the significance of the dynamic changes in F-actin within the fertilized egg has been largely unclear.
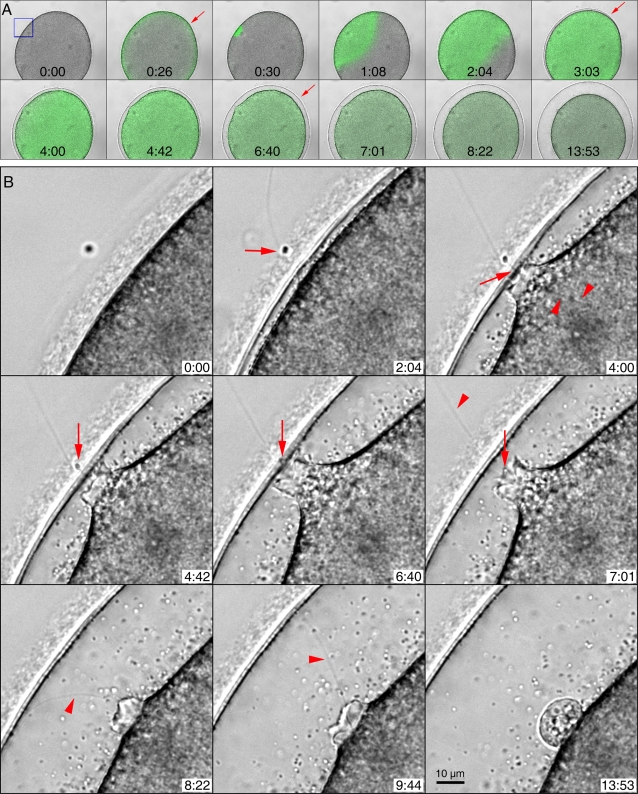
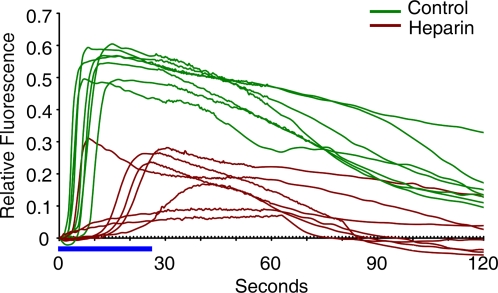
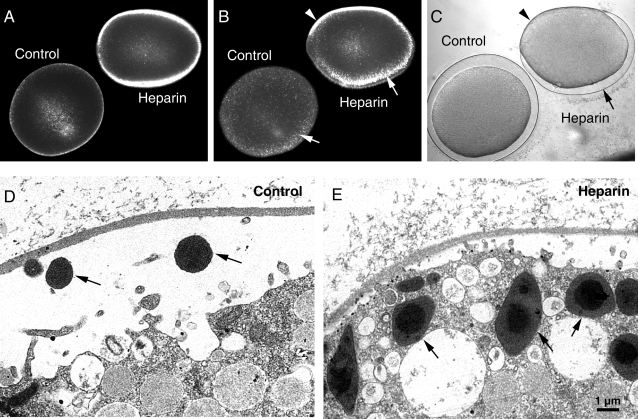
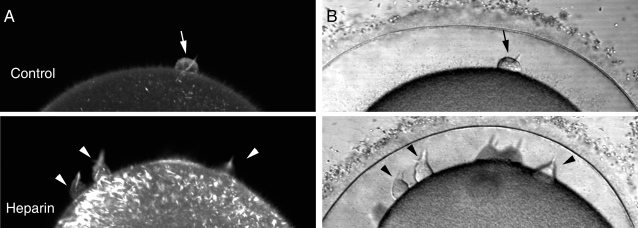
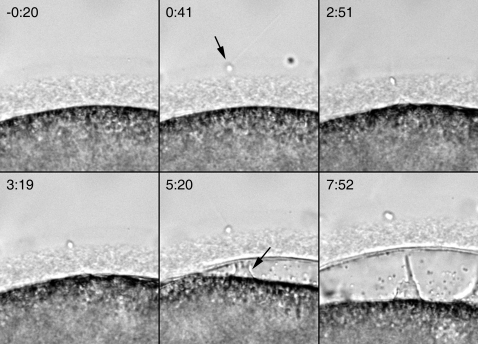
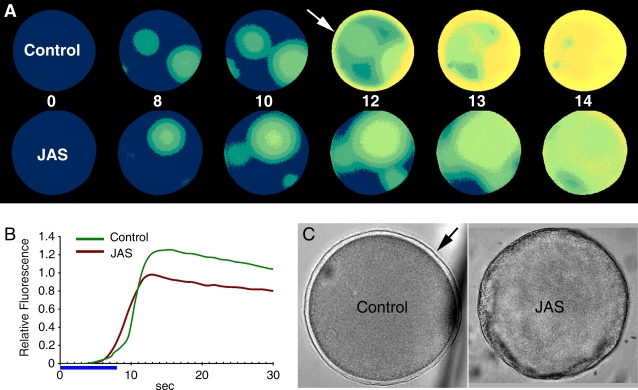
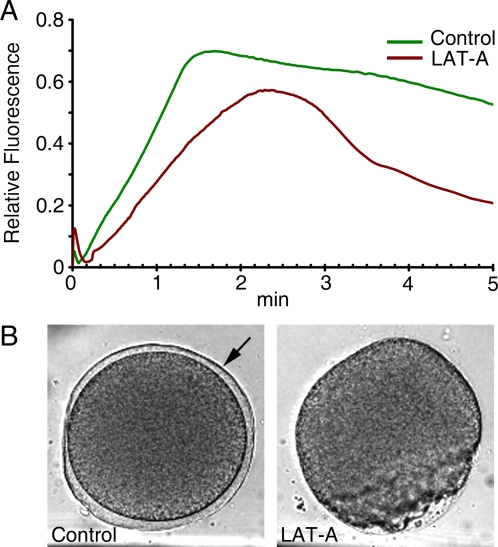
Methodology/principal findings: We have measured changes in intracellular Ca2+ signals and F-actin structures during fertilization. We also report the unexpected observation that the conventional antagonist of the InsP(3) receptor, heparin, hyperpolymerizes the cortical actin cytoskeleton in postmeiotic eggs. Using heparin and other pharmacological agents that either hypo- or hyperpolymerize the cortical actin, we demonstrate that nearly all aspects of the fertilization process are profoundly affected by the dynamic restructuring of the egg cortical actin cytoskeleton.
Conclusions/significance: Our findings identify important roles for subplasmalemmal actin fibers in the process of sperm-egg interaction and in the subsequent events related to fertilization: the generation of Ca2+ signals, sperm penetration, cortical granule exocytosis, and the block to polyspermy.
Conflict of interest statement
Figures


References
-
- Dan JC. Studies on the acrosome. II Acrosome reaction in starfish spermatozoa. Biol Bull. 1954;107:203–218.
-
- Dale B, Dan-Sohkawa M, De Santis A, Hoshi M. Fertilization of the starfish Astropecten aurantiacus. Exp Cell Res. 1981;132:505–510. - PubMed
-
- Ikadai H, Hoshi M. Biochemical studies on the acrosome reaction of the starfish Asterias amurensis. I. Factors participating in the acrosome reaction. Dev Growth Diff. 1982;23:73–80. - PubMed
-
- Kyozuka K, Osanai K. Fertilization cone formation in starfish oocytes: the role of the egg cortex actin microfilaments in sperm incorporation. Gamete Res. 1988;20:275–285. - PubMed
-
- Miyazaki S, Hirai S. Fast polyspermy block and activation potential. Correlated changes during oocyte maturation of a starfish. Dev Biol. 1979;70:327–340. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
