

Recruitment of the complete hTREX complex is required for Kaposi's sarcoma-associated herpesvirus intronless mRNA nuclear export and virus replication
- PMID: 18974867
- PMCID: PMC2569588
- DOI: 10.1371/journal.ppat.1000194
Recruitment of the complete hTREX complex is required for Kaposi's sarcoma-associated herpesvirus intronless mRNA nuclear export and virus replication
Abstract
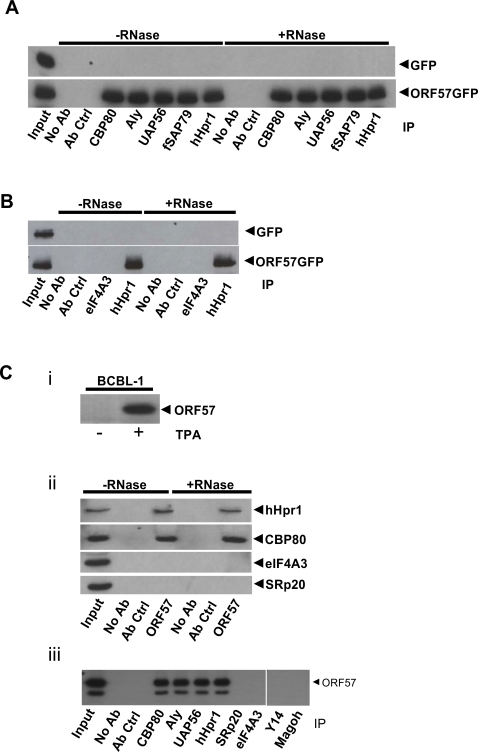
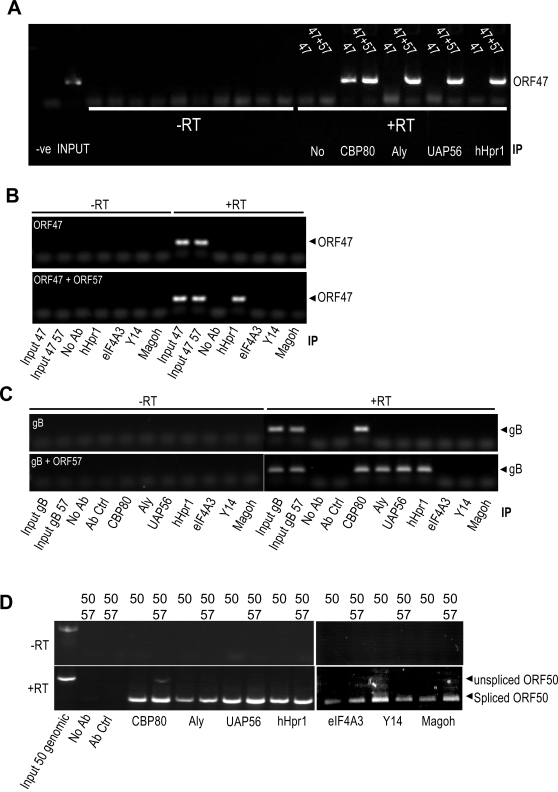
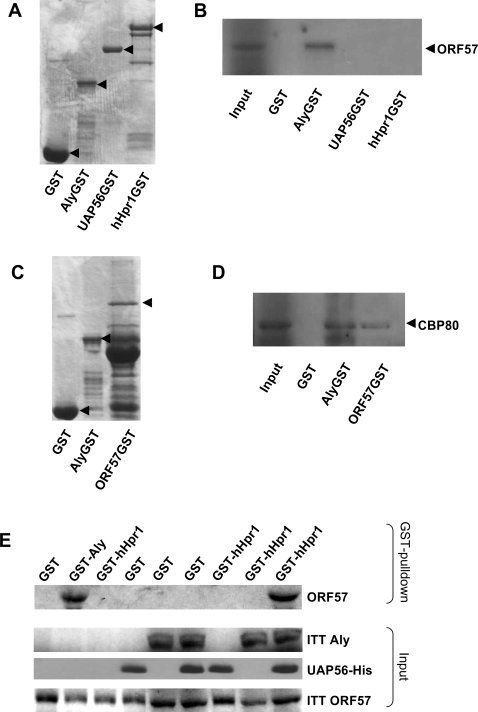
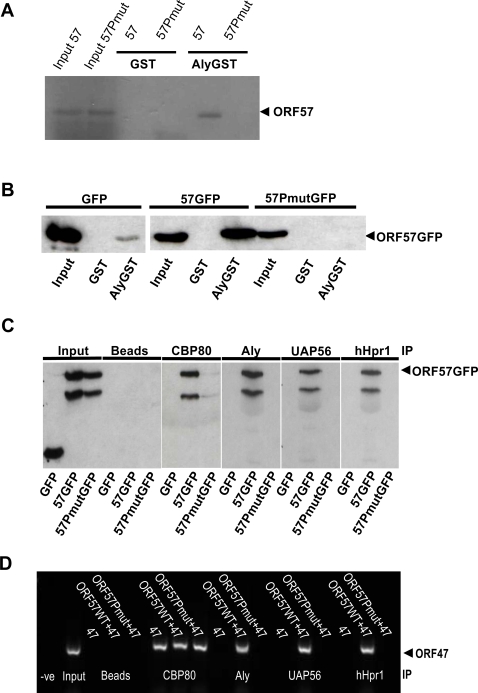
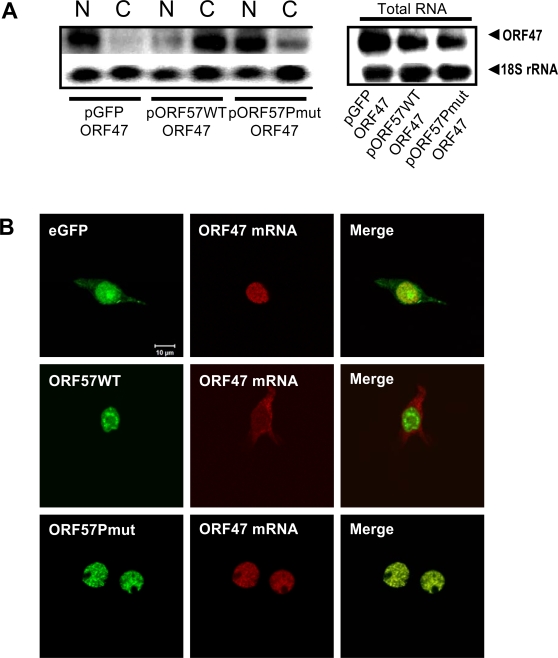
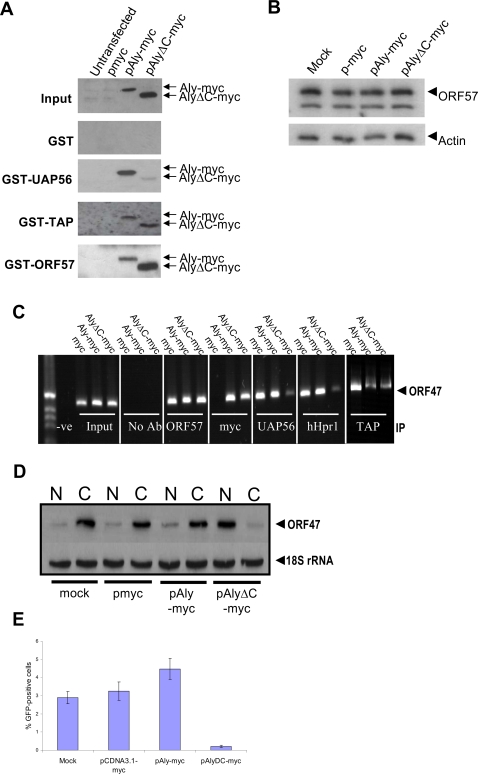
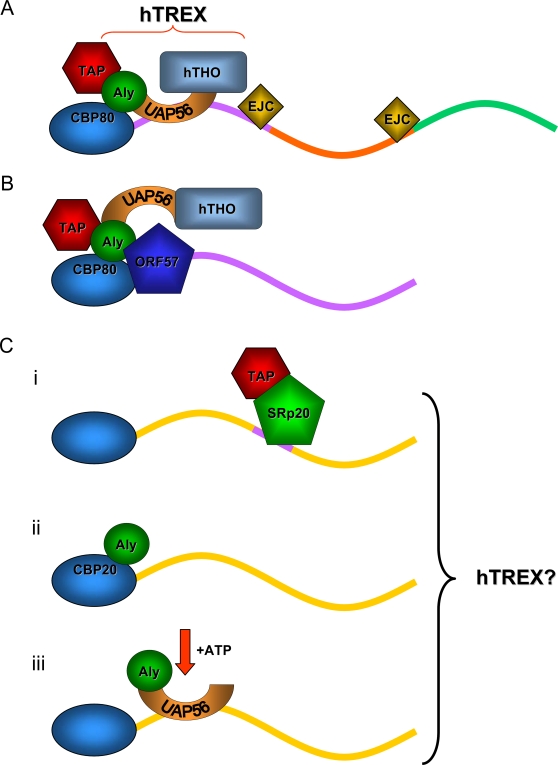
A cellular pre-mRNA undergoes various post-transcriptional processing events, including capping, splicing and polyadenylation prior to nuclear export. Splicing is particularly important for mRNA nuclear export as two distinct multi-protein complexes, known as human TREX (hTREX) and the exon-junction complex (EJC), are recruited to the mRNA in a splicing-dependent manner. In contrast, a number of Kaposi's sarcoma-associated herpesvirus (KSHV) lytic mRNAs lack introns and are exported by the virus-encoded ORF57 protein. Herein we show that ORF57 binds to intronless viral mRNAs and functions to recruit the complete hTREX complex, but not the EJC, in order assemble an export component viral ribonucleoprotein particle (vRNP). The formation of this vRNP is mediated by a direct interaction between ORF57 and the hTREX export adapter protein, Aly. Aly in turn interacts directly with the DEAD-box protein UAP56, which functions as a bridge to recruit the remaining hTREX proteins to the complex. Moreover, we show that a point mutation in ORF57 which disrupts the ORF57-Aly interaction leads to a failure in the ORF57-mediated recruitment of the entire hTREX complex to the intronless viral mRNA and inhibits the mRNAs subsequent nuclear export and virus replication. Furthermore, we have utilised a trans-dominant Aly mutant to prevent the assembly of the complete ORF57-hTREX complex; this results in a vRNP consisting of viral mRNA bound to ORF57, Aly and the nuclear export factor, TAP. Strikingly, although both the export adapter Aly and the export factor TAP were present on the viral mRNP, a dramatic decrease in intronless viral mRNA export and virus replication was observed in the absence of the remaining hTREX components (UAP56 and hTHO-complex). Together, these data provide the first direct evidence that the complete hTREX complex is essential for the export of KSHV intronless mRNAs and infectious virus production.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Bentley DL. Rules of engagement: co-transcriptional recruitment of pre-mRNA processing factors. Curr Opin Cell Biol. 2005;17:251–256. - PubMed
-
- Erkmann JA, Kutay U. Nuclear export of mRNA: from the site of transcription to the cytoplasm. Exp Cell Res. 2004;296:12–20. - PubMed
-
- Moore MJ. From birth to death: the complex lives of eukaryotic mRNAs. Science. 2005;309:1514–1518. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
