

Stripes and belly-spots -- a review of pigment cell morphogenesis in vertebrates
- PMID: 18977309
- PMCID: PMC2744437
- DOI: 10.1016/j.semcdb.2008.10.001
Stripes and belly-spots -- a review of pigment cell morphogenesis in vertebrates
Abstract
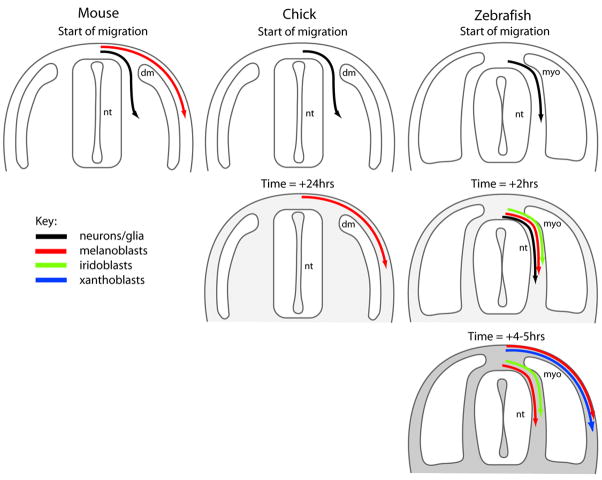
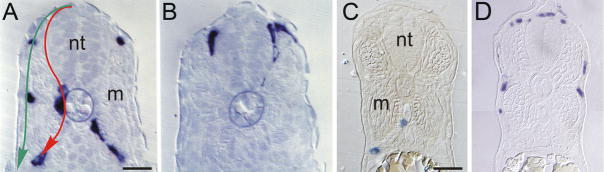
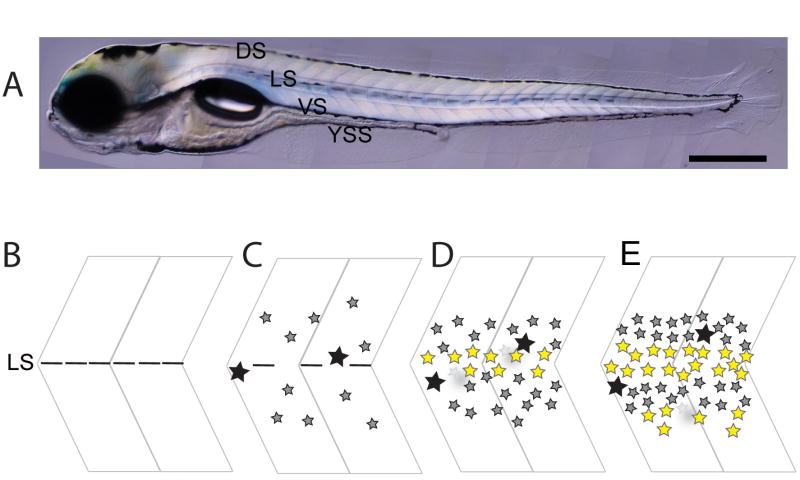
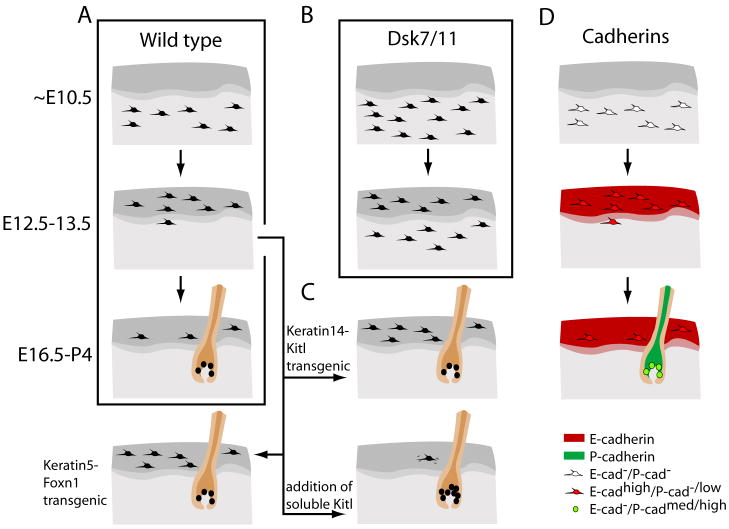
Pigment patterns in the integument have long-attracted attention from both scientists and non-scientists alike since their natural attractiveness combines with their excellence as models for the general problem of pattern formation. Pigment cells are formed from the neural crest and must migrate to reach their final locations. In this review, we focus on our current understanding of mechanisms underlying the control of pigment cell migration and patterning in diverse vertebrates. The model systems discussed here - chick, mouse, and zebrafish - each provide unique insights into the major morphogenetic events driving pigment pattern formation. In birds and mammals, melanoblasts must be specified before they can migrate on the dorsolateral pathway. Transmembrane receptors involved in guiding them onto this route include EphB2 and Ednrb2 in chick, and Kit in mouse. Terminal migration depends, in part, upon extracellular matrix reorganization by ADAMTS20. Invasion of the ectoderm, especially into the feather germ and hair follicles, requires specific signals that are beginning to be characterized. We summarize our current understanding of the mechanisms regulating melanoblast number and organization in the epidermis. We note the apparent differences in pigment pattern formation in poikilothermic vertebrates when compared with birds and mammals. With more pigment cell types, migration pathways are more complex and largely unexplored; nevertheless, a role for Kit signaling in melanophore migration is clear and indicates that at least some patterning mechanisms may be highly conserved. We summarize the multiple factors thought to contribute to zebrafish embryonic pigment pattern formation, highlighting a recent study identifying Sdf1a as one factor crucial for regulation of melanophore positioning. Finally, we discuss the mechanisms generating a second, metamorphic pigment pattern in adult fish, emphasizing recent studies strengthening the evidence that undifferentiated progenitor cells play a major role in generating adult pigment cells.
Figures

References
-
- Le Douarin NM, Kalcheim C. The Neural Crest. 2. Cambridge: Cambridge University Press; 1999.
-
- Fox MH. Analysis of some phases of melanoblast migration in barred plymouth rock embryos. Physiol Zool. 1949;22:1–22. - PubMed
-
- Ris H. An experimental study of the origin of melanophores in birds. Physiol Zool. 1941;14:48–66.
-
- Watterson RL. The morphogenesis of down feathers with special reference to the developmental history of melanophores. Physiol Zool. 1942;15:234–259.
-
- Willier BH, Rawles ME. The control of feather color pattern by melanophores grafted from one embryo to another of a different breed of fowl. Physiol Zool. 1940;13:177–202.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
