

Kinetic and X-ray structural evidence for negative cooperativity in substrate binding to nicotinate mononucleotide adenylyltransferase (NMAT) from Bacillus anthracis
- PMID: 18977360
- PMCID: PMC2654404
- DOI: 10.1016/j.jmb.2008.10.037
Kinetic and X-ray structural evidence for negative cooperativity in substrate binding to nicotinate mononucleotide adenylyltransferase (NMAT) from Bacillus anthracis
Abstract
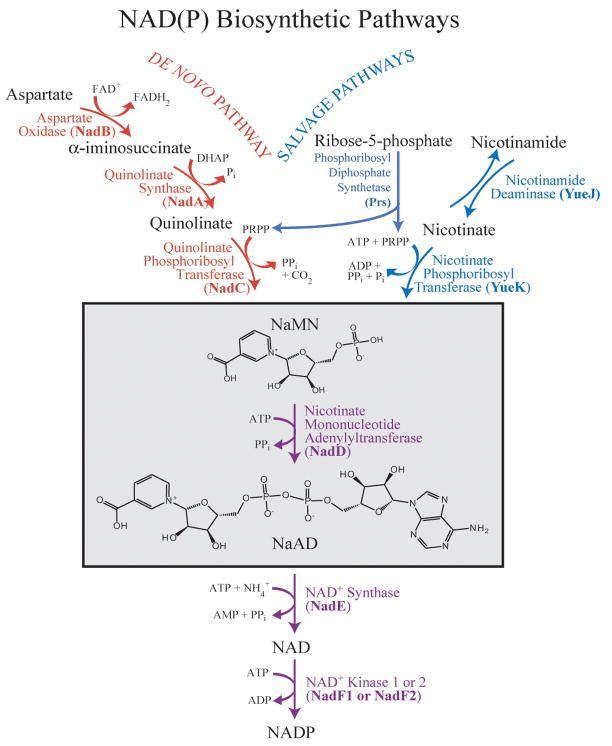
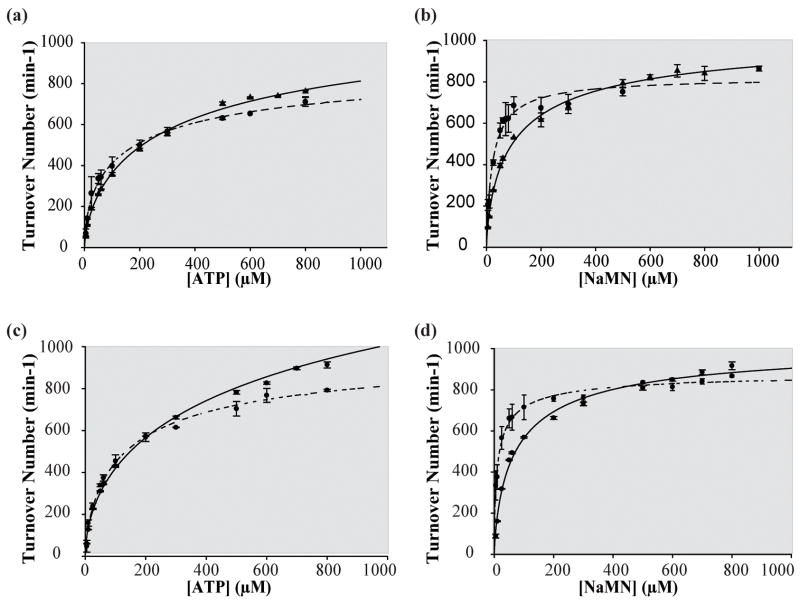
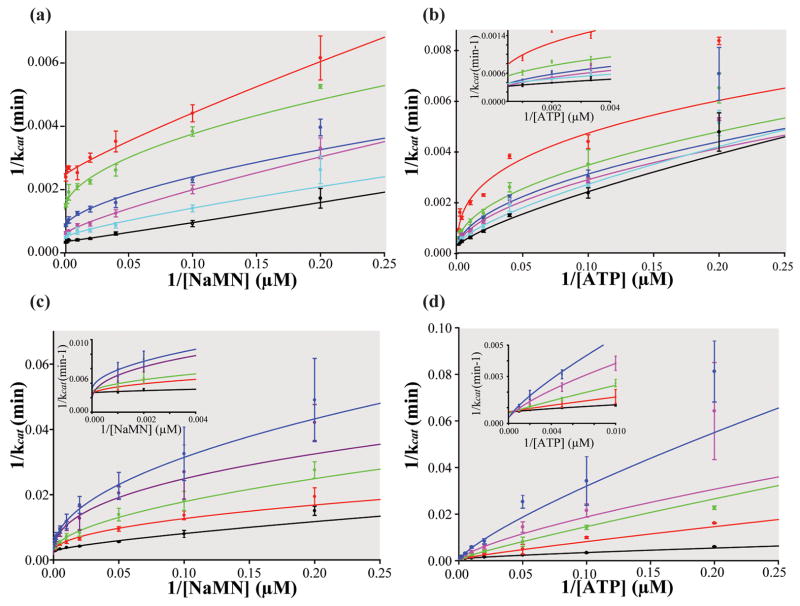
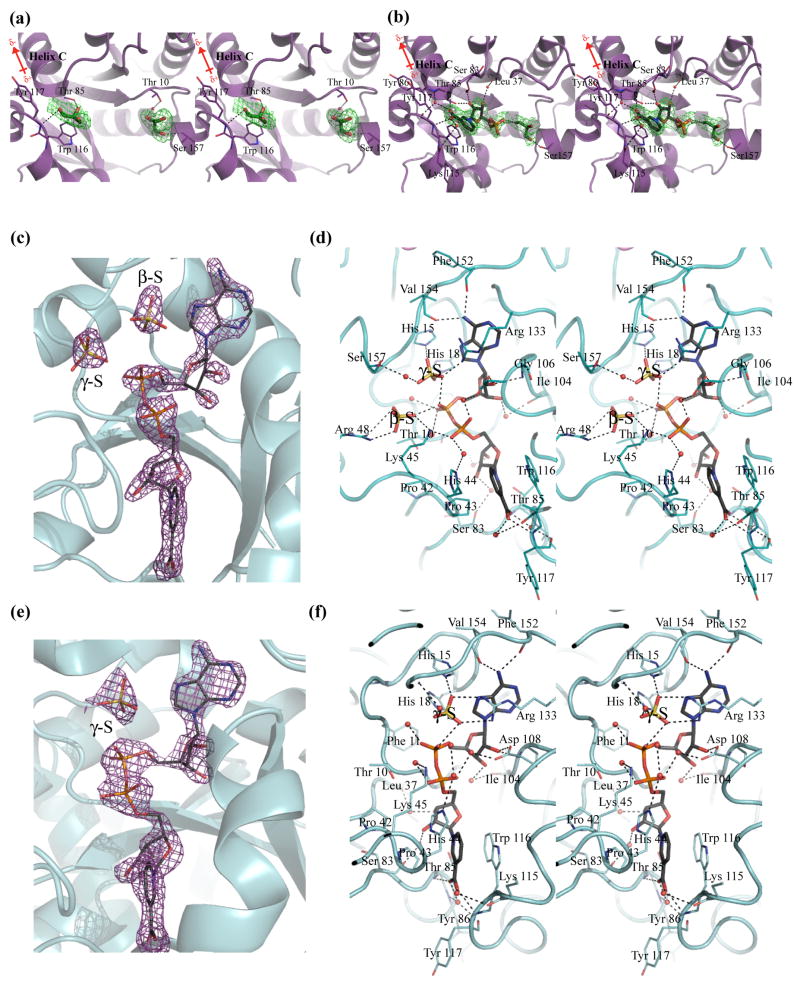
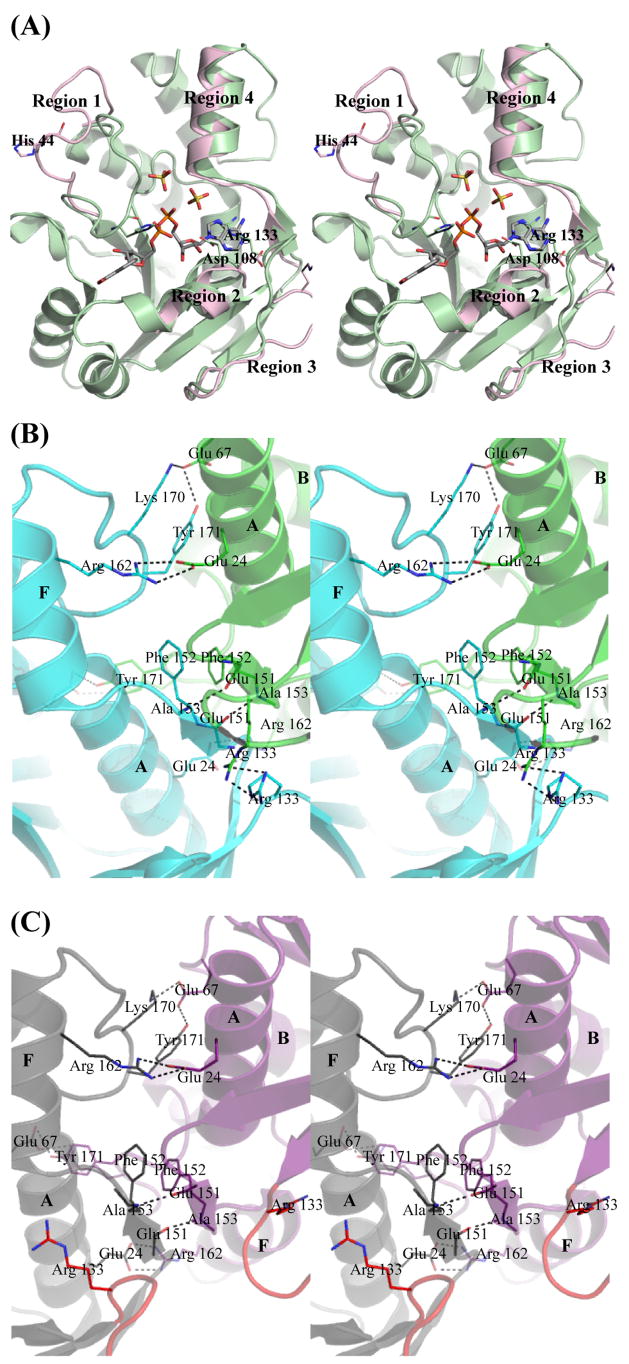
Biosynthesis of NAD(P) in bacteria occurs either de novo or through one of the salvage pathways that converge at the point where the reaction of nicotinate mononucleotide (NaMN) with ATP is coupled to the formation of nicotinate adenine dinucleotide (NaAD) and inorganic pyrophosphate. This reaction is catalyzed by nicotinate mononucleotide adenylyltransferase (NMAT), which is essential for bacterial growth, making it an attractive drug target for the development of new antibiotics. Steady-state kinetic and direct binding studies on NMAT from Bacillus anthracis suggest a random sequential Bi-Bi kinetic mechanism. Interestingly, the interactions of NaMN and ATP with NMAT were observed to exhibit negative cooperativity, i.e. Hill coefficients <1.0. Negative cooperativity in binding is supported by the results of X-ray crystallographic studies. X-ray structures of the B. anthracis NMAT apoenzyme, and the NaMN- and NaAD-bound complexes were determined to resolutions of 2.50 A, 2.60 A and 1.75 A, respectively. The X-ray structure of the NMAT-NaMN complex revealed only one NaMN molecule bound in the biological dimer, supporting negative cooperativity in substrate binding. The kinetic, direct-binding, and X-ray structural studies support a model in which the binding affinity of substrates to the first monomer of NMAT is stronger than that to the second, and analysis of the three X-ray structures reveals significant conformational changes of NMAT along the enzymatic reaction coordinate. The negative cooperativity observed in B. anthracis NMAT substrate binding is a unique property that has not been observed in other prokaryotic NMAT enzymes. We propose that regulation of the NAD(P) biosynthetic pathway may occur, in part, at the reaction catalyzed by NMAT.
Figures


References
-
- Kalamas AG. Anthrax. Anethesiology Clinics of North America. 2004:22. - PubMed
-
- Atlas RM. Bioterriorism: From threat to reality. Annu Rev Microbiol. 2002;56:167–185. - PubMed
-
- Athamna A, Athamna M, Abu-Rashed N, Medlej B, Bast DJ, Rubinstein E. Selection of Bacillus anthracis isolates resistant to antibiotics. J Antimicrob Chemother. 2004;54:424–428. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
