

Asymmetric division of activated latently infected cells may explain the decay kinetics of the HIV-1 latent reservoir and intermittent viral blips
- PMID: 18977369
- PMCID: PMC2657607
- DOI: 10.1016/j.mbs.2008.10.006
Asymmetric division of activated latently infected cells may explain the decay kinetics of the HIV-1 latent reservoir and intermittent viral blips
Abstract
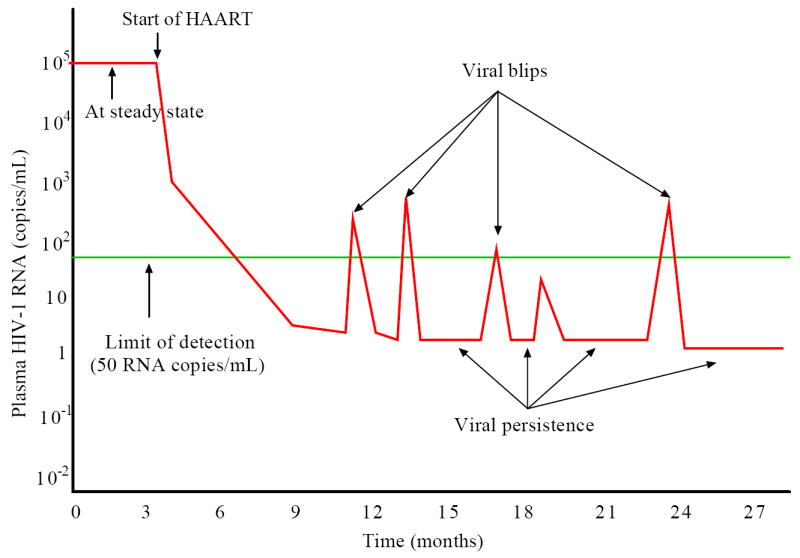
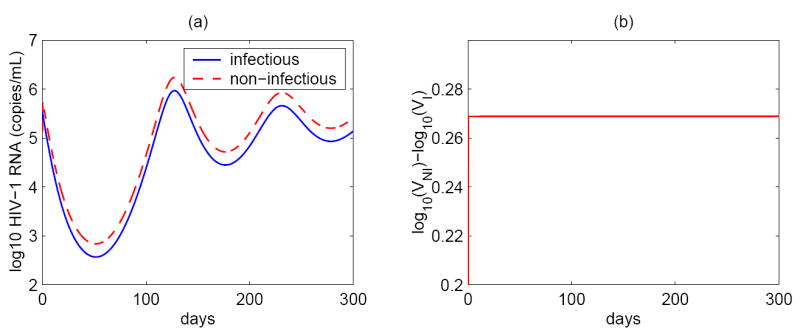
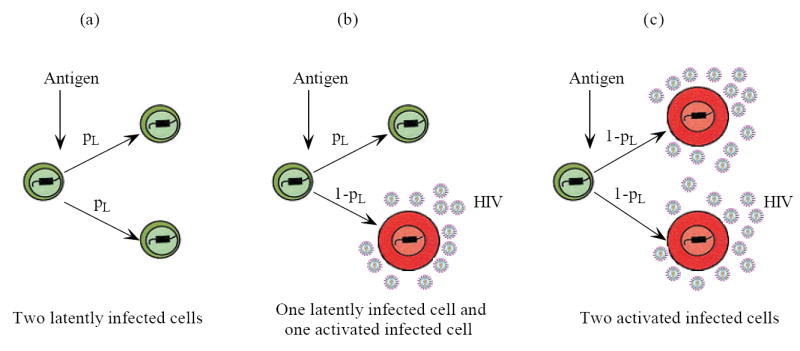
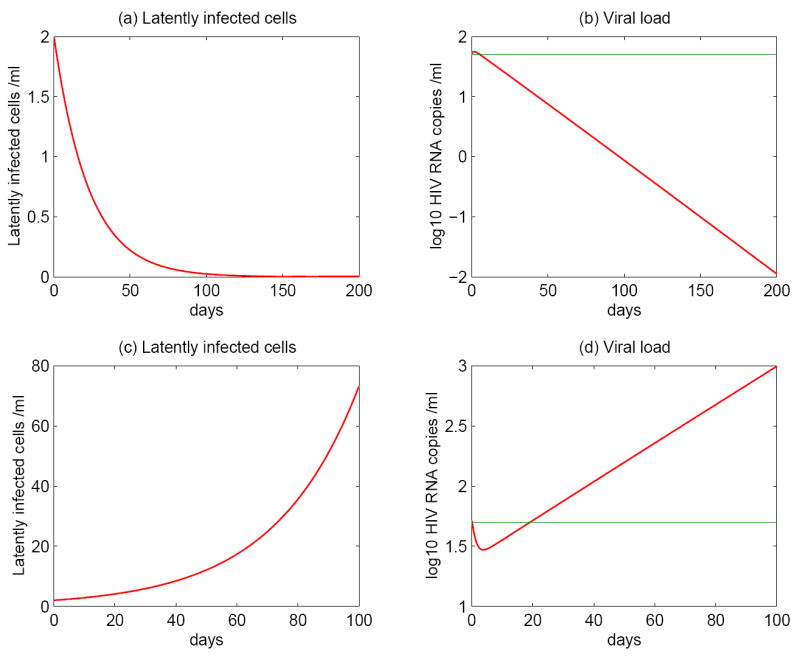
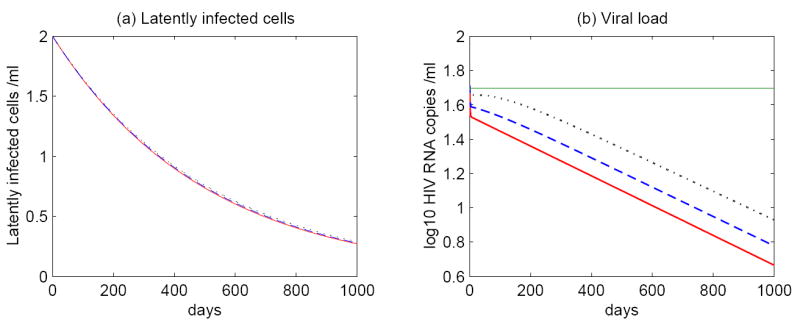
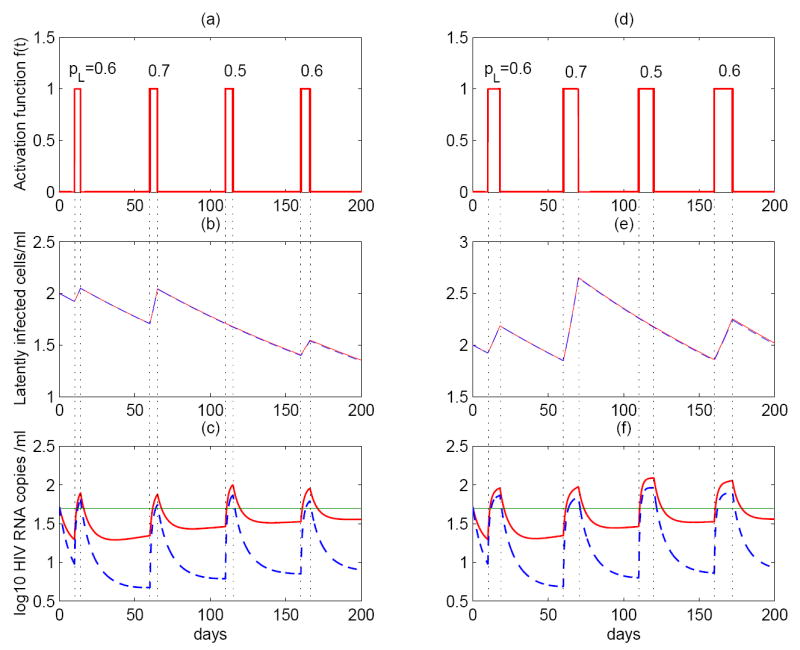
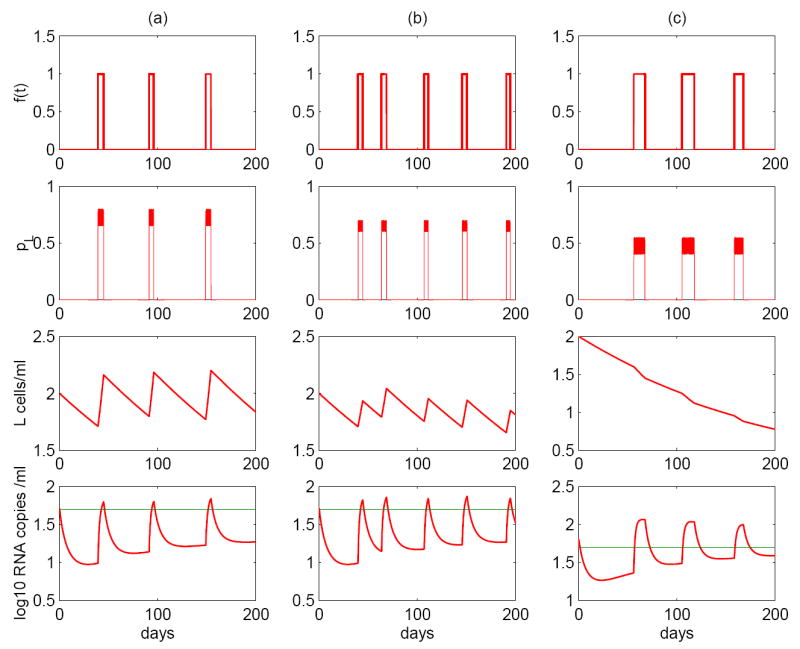
Most HIV-infected patients when treated with combination antiretroviral therapy achieve viral loads that are below the current limit of detection of standard assays after a few months. Despite this, virus eradication from the host has not been achieved. Latent, replication-competent HIV-1 can generally be identified in resting memory CD4(+) T cells in patients with "undetectable" viral loads. Turnover of these cells is extremely slow but virus can be released from the latent reservoir quickly upon cessation of therapy. In addition, a number of patients experience transient episodes of viremia, or HIV-1 blips, even with suppression of the viral load to below the limit of detection for many years. The mechanisms underlying the slow decay of the latent reservoir and the occurrence of intermittent viral blips have not been fully elucidated. In this study, we address these two issues by developing a mathematical model that explores a hypothesis about latently infected cell activation. We propose that asymmetric division of latently infected cells upon sporadic antigen encounter may both replenish the latent reservoir and generate intermittent viral blips. Interestingly, we show that occasional replenishment of the latent reservoir induced by reactivation of latently infected cells may reconcile the differences between the divergent estimates of the half-life of the latent reservoir in the literature.
Figures

Similar articles
-
Modeling HIV persistence, the latent reservoir, and viral blips.J Theor Biol. 2009 Sep 21;260(2):308-31. doi: 10.1016/j.jtbi.2009.06.011. Epub 2009 Jun 17. J Theor Biol. 2009. PMID: 19539630 Free PMC article. Review.
-
Modeling latently infected cell activation: viral and latent reservoir persistence, and viral blips in HIV-infected patients on potent therapy.PLoS Comput Biol. 2009 Oct;5(10):e1000533. doi: 10.1371/journal.pcbi.1000533. Epub 2009 Oct 16. PLoS Comput Biol. 2009. PMID: 19834532 Free PMC article.
-
Stochastic population switch may explain the latent reservoir stability and intermittent viral blips in HIV patients on suppressive therapy.J Theor Biol. 2014 Nov 7;360:137-148. doi: 10.1016/j.jtbi.2014.06.042. Epub 2014 Jul 10. J Theor Biol. 2014. PMID: 25016044
-
Transient viremia, plasma viral load, and reservoir replenishment in HIV-infected patients on antiretroviral therapy.J Acquir Immune Defic Syndr. 2007 Aug 15;45(5):483-93. doi: 10.1097/QAI.0b013e3180654836. J Acquir Immune Defic Syndr. 2007. PMID: 17496565 Free PMC article.
-
Reservoirs for HIV-1: mechanisms for viral persistence in the presence of antiviral immune responses and antiretroviral therapy.Annu Rev Immunol. 2000;18:665-708. doi: 10.1146/annurev.immunol.18.1.665. Annu Rev Immunol. 2000. PMID: 10837072 Review.
Cited by
-
Quantifying the treatment efficacy of reverse transcriptase inhibitors: new analyses of clinical data based on within-host modeling.BMC Public Health. 2009 Nov 18;9 Suppl 1(Suppl 1):S11. doi: 10.1186/1471-2458-9-S1-S11. BMC Public Health. 2009. PMID: 19922681 Free PMC article. Clinical Trial.
-
Modeling HIV persistence, the latent reservoir, and viral blips.J Theor Biol. 2009 Sep 21;260(2):308-31. doi: 10.1016/j.jtbi.2009.06.011. Epub 2009 Jun 17. J Theor Biol. 2009. PMID: 19539630 Free PMC article. Review.
-
Mathematical Models of HIV Latency.Curr Top Microbiol Immunol. 2018;417:131-156. doi: 10.1007/82_2017_77. Curr Top Microbiol Immunol. 2018. PMID: 29164341 Free PMC article. Review.
-
Conflict and accord of optimal treatment strategies for HIV infection within and between hosts.Math Biosci. 2019 Mar;309:107-117. doi: 10.1016/j.mbs.2019.01.007. Epub 2019 Jan 24. Math Biosci. 2019. PMID: 30684516 Free PMC article.
-
Modeling dynamics of acute HIV infection incorporating density-dependent cell death and multiplicity of infection.PLoS Comput Biol. 2024 Jun 7;20(6):e1012129. doi: 10.1371/journal.pcbi.1012129. eCollection 2024 Jun. PLoS Comput Biol. 2024. PMID: 38848426 Free PMC article.
References
-
- Collier AC, Coombs RW, Schoenfeld DA, et al. Treatment of human immunodeficiency virus infection with saquinavir, zidovudine, and zalcitabine. N Engl J Med. 1996;334:1011. - PubMed
-
- Dornadula G, Zhang H, VanUitert B, et al. Residual HIV-1 RNA in blood plasma of patients taking suppressive highly active antiretroviral therapy. JAMA. 1999;282:1627. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
