

Genetic interactions of smc, ftsK, and parB genes in Streptomyces coelicolor and their developmental genome segregation phenotypes
- PMID: 18978061
- PMCID: PMC2612423
- DOI: 10.1128/JB.00858-08
Genetic interactions of smc, ftsK, and parB genes in Streptomyces coelicolor and their developmental genome segregation phenotypes
Abstract
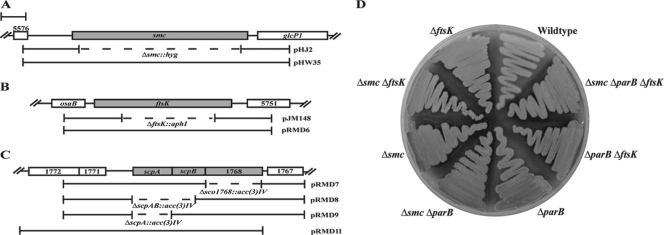
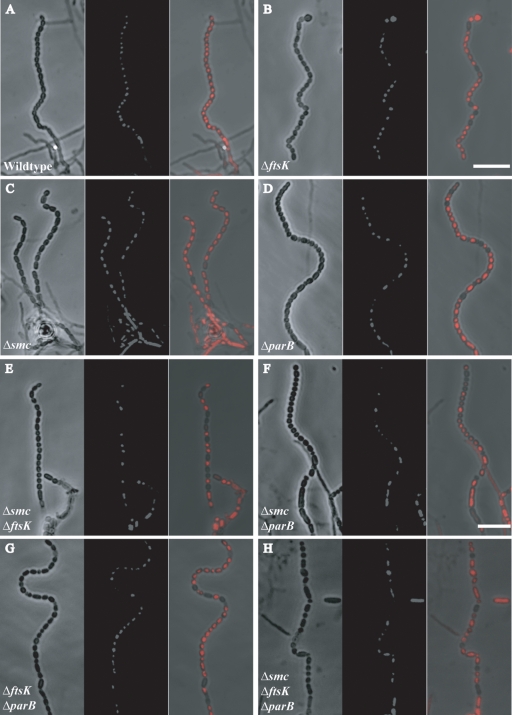
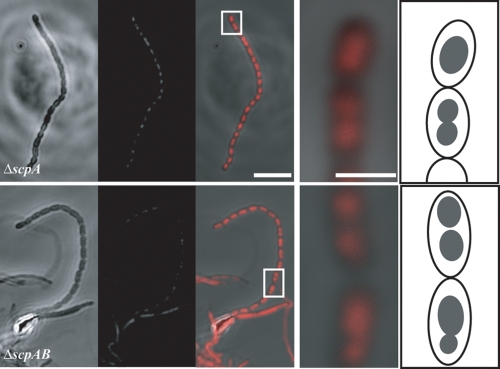
The mechanisms by which chromosomes condense and segregate during developmentally regulated cell division are of interest for Streptomyces coelicolor, a sporulating, filamentous bacterium with a large, linear genome. These processes coordinately occur as many septa synchronously form in syncytial aerial hyphae such that prespore compartments accurately receive chromosome copies. Our genetic approach analyzed mutants for ftsK, smc, and parB. DNA motor protein FtsK/SpoIIIE coordinates chromosome segregation with septum closure in rod-shaped bacteria. SMC (structural maintenance of chromosomes) participates in condensation and organization of the nucleoid. ParB/Spo0J partitions the origin of replication using a nucleoprotein complex, assembled at a centromere-like sequence. Consistent with previous work, we show that an ftsK-null mutant produces anucleate spores at the same frequency as the wild-type strain (0.8%). We report that the smc and ftsK deletion-insertion mutants (ftsK' truncation allele) have developmental segregation defects (7% and 15% anucleate spores, respectively). By use of these latter mutants, viable double and triple mutants were isolated in all combinations with a previously described parB-null mutant (12% anucleate spores). parB and smc were in separate segregation pathways; the loss of both exacerbates the segregation defect (24% anucleate spores). For a triple mutant, deletion of the region encoding the FtsK motor domain and one transmembrane segment partially alleviates the segregation defect of the smc parB mutant (10% anucleate spores). Considerable redundancy must exist in this filamentous organism because segregation of some genomic material occurs 90% of the time during development in the absence of three functions with only a fourfold loss of spore viability. Furthermore, we report that scpA and scpAB mutants (encoding SMC-associated proteins) have spore nucleoid organization defects. Finally, FtsK-enhanced green fluorescent protein (EGFP) localized as bands or foci between incipient nucleoids, while SMC-EGFP foci were not uniformly positioned along aerial hyphae, nor were they associated with every condensing nucleoid.
Figures

Similar articles
-
SMC protein-dependent chromosome condensation during aerial hyphal development in Streptomyces.J Bacteriol. 2009 Jan;191(1):310-9. doi: 10.1128/JB.00513-08. Epub 2008 Oct 17. J Bacteriol. 2009. PMID: 18931116 Free PMC article.
-
Role of an FtsK-like protein in genetic stability in Streptomyces coelicolor A3(2).J Bacteriol. 2007 Mar;189(6):2310-8. doi: 10.1128/JB.01660-06. Epub 2007 Jan 5. J Bacteriol. 2007. PMID: 17209017 Free PMC article.
-
Early onset of septal FtsK localization allows for efficient DNA segregation in SMC-deleted Corynebacterium glutamicum strains.mBio. 2025 Mar 12;16(3):e0285924. doi: 10.1128/mbio.02859-24. Epub 2025 Jan 28. mBio. 2025. PMID: 39873485 Free PMC article.
-
[Chromosome segregation and cell division during the growth and differentiation of Streptomyces].Postepy Hig Med Dosw (Online). 2007 Oct 12;61:565-75. Postepy Hig Med Dosw (Online). 2007. PMID: 17971760 Review. Polish.
-
Building bridges within the bacterial chromosome.Trends Genet. 2015 Mar;31(3):164-73. doi: 10.1016/j.tig.2015.01.003. Epub 2015 Feb 11. Trends Genet. 2015. PMID: 25682183 Review.
Cited by
-
Membrane-associated DNA transport machines.Cold Spring Harb Perspect Biol. 2010 Jul;2(7):a000406. doi: 10.1101/cshperspect.a000406. Epub 2010 Jun 23. Cold Spring Harb Perspect Biol. 2010. PMID: 20573715 Free PMC article. Review.
-
A septal chromosome segregator protein evolved into a conjugative DNA-translocator protein.Mob Genet Elements. 2011 Sep;1(3):225-229. doi: 10.4161/mge.1.3.18066. Epub 2011 Sep 1. Mob Genet Elements. 2011. PMID: 22479692 Free PMC article.
-
Taxonomy, Physiology, and Natural Products of Actinobacteria.Microbiol Mol Biol Rev. 2015 Nov 25;80(1):1-43. doi: 10.1128/MMBR.00019-15. Print 2016 Mar. Microbiol Mol Biol Rev. 2015. PMID: 26609051 Free PMC article. Review.
-
Structure-function relationships of two paralogous single-stranded DNA-binding proteins from Streptomyces coelicolor: implication of SsbB in chromosome segregation during sporulation.Nucleic Acids Res. 2013 Apr 1;41(6):3659-72. doi: 10.1093/nar/gkt050. Epub 2013 Feb 7. Nucleic Acids Res. 2013. PMID: 23393191 Free PMC article.
-
Quantitative Proteome and Phosphoproteome Analyses of Streptomyces coelicolor Reveal Proteins and Phosphoproteins Modulating Differentiation and Secondary Metabolism.Mol Cell Proteomics. 2018 Aug;17(8):1591-1611. doi: 10.1074/mcp.RA117.000515. Epub 2018 May 21. Mol Cell Proteomics. 2018. PMID: 29784711 Free PMC article.
References
-
- Ausmees, N., H. Wahlstedt, S. Bagchi, M. A. Elliot, M. J. Buttner, and K. Flardh. 2007. SmeA, a small membrane protein with multiple functions in Streptomyces sporulation including targeting of a SpoIIIE/FtsK-like protein to cell division septa. Mol. Microbiol. 651458-1473. - PubMed
-
- Barre, F. X. 2007. FtsK and SpoIIIE: the tale of the conserved tails. Mol. Microbiol. 661051-1055. - PubMed
-
- Bentley, S. D., K. F. Chater, A. M. Cerdeno-Tarraga, G. L. Challis, N. R. Thomson, K. D. James, D. E. Harris, M. A. Quail, H. Kieser, D. Harper, A. Bateman, S. Brown, G. Chandra, C. W. Chen, M. Collins, A. Cronin, A. Fraser, A. Goble, J. Hidalgo, T. Hornsby, S. Howarth, C. H. Huang, T. Kieser, L. Larke, L. Murphy, K. Oliver, S. O'Neil, E. Rabbinowitsch, M. A. Rajandream, K. Rutherford, S. Rutter, K. Seeger, D. Saunders, S. Sharp, R. Squares, S. Squares, K. Taylor, T. Warren, A. Wietzorrek, J. Woodward, B. G. Barrell, J. Parkhill, and D. A. Hopwood. 2002. Complete genome sequence of the model actinomycete Streptomyces coelicolor A3(2). Nature 417141-147. - PubMed
-
- Bierman, M., R. Logan, K. O'Brien, E. T. Seno, R. N. Rao, and B. E. Schoner. 1992. Plasmid cloning vectors for the conjugal transfer of DNA from Escherichia coli to Streptomyces spp. Gene 11643-49. - PubMed
-
- Bignell, C., and C. M. Thomas. 2001. The bacterial ParA-ParB partitioning proteins. J. Biotechnol. 911-34. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
