

Layer, column and cell-type specific genetic manipulation in mouse barrel cortex
- PMID: 18982108
- PMCID: PMC2570061
- DOI: 10.3389/neuro.01.001.2008
Layer, column and cell-type specific genetic manipulation in mouse barrel cortex
Abstract
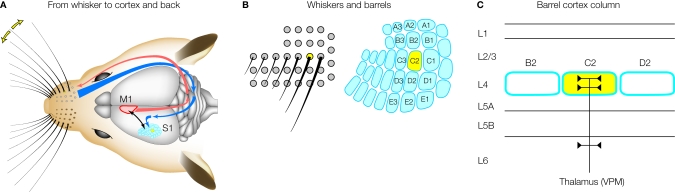
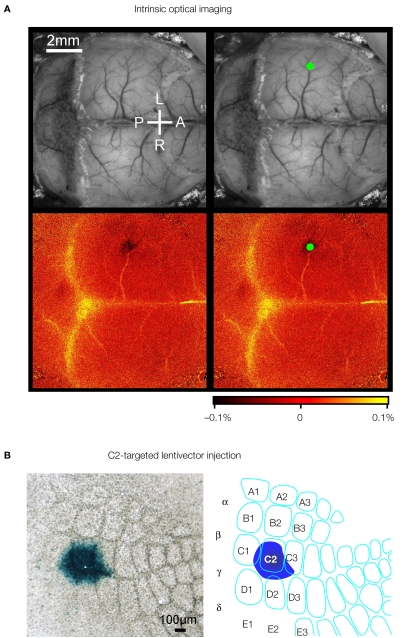
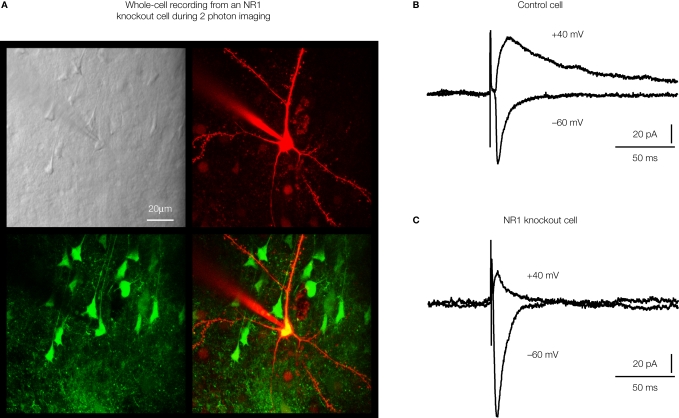
Sensory information is processed in distributed neuronal networks connected by intricate synaptic circuits. Studies of the rodent brain can provide insight into synaptic mechanisms of sensory perception and associative learning. In particular, the mouse whisker sensorimotor system has recently begun to be investigated through combinations of imaging and electrophysiology, providing data correlating neural activity with behaviour. In order to go beyond such correlative studies and to pinpoint the contributions of individual genes to brain function, it is critical to make highly controlled and specific manipulations. Here, we review recent progress towards genetic manipulation of targeted genes in specific neuronal cell types located in a selected cortical layer of a well-defined cortical column of mouse barrel cortex. The unprecedented precision of such genetic manipulation within highly specific neural circuits may contribute significantly to progress in understanding the molecular and synaptic determinants of simple forms of sensory perception and associative learning.
Keywords: NMDA receptors; barrel cortex; genetic manipulation; lentivirus.
Figures


References
LinkOut - more resources
Full Text Sources