

Differential activities of cellular and viral macro domain proteins in binding of ADP-ribose metabolites
- PMID: 18983849
- PMCID: PMC7094737
- DOI: 10.1016/j.jmb.2008.10.045
Differential activities of cellular and viral macro domain proteins in binding of ADP-ribose metabolites
Abstract
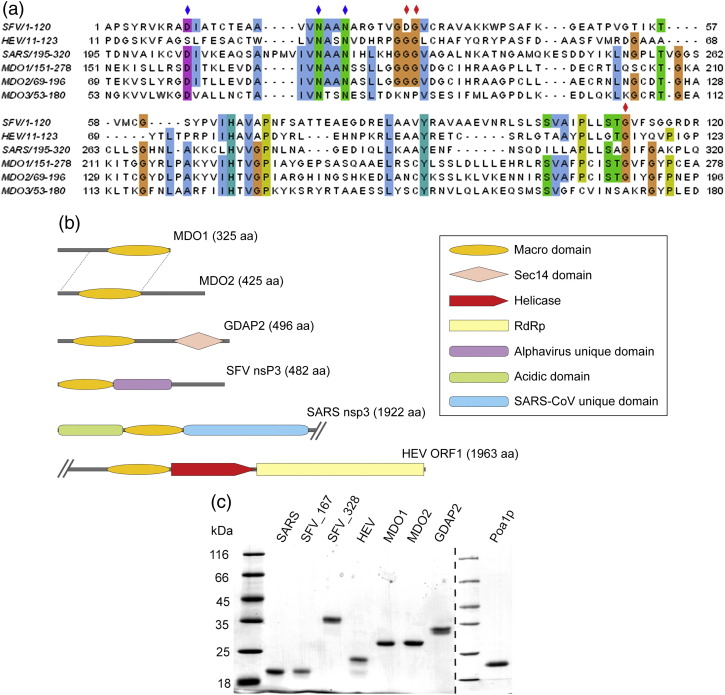
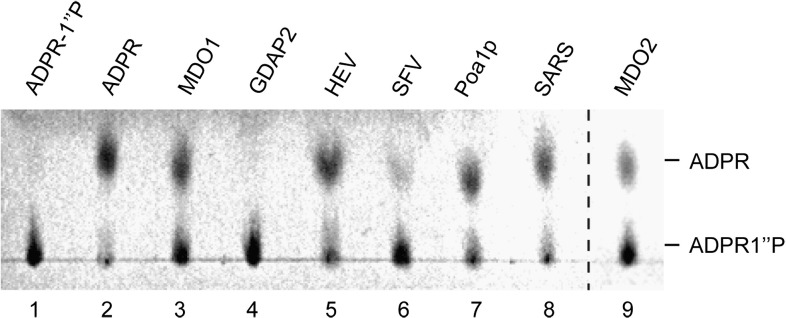
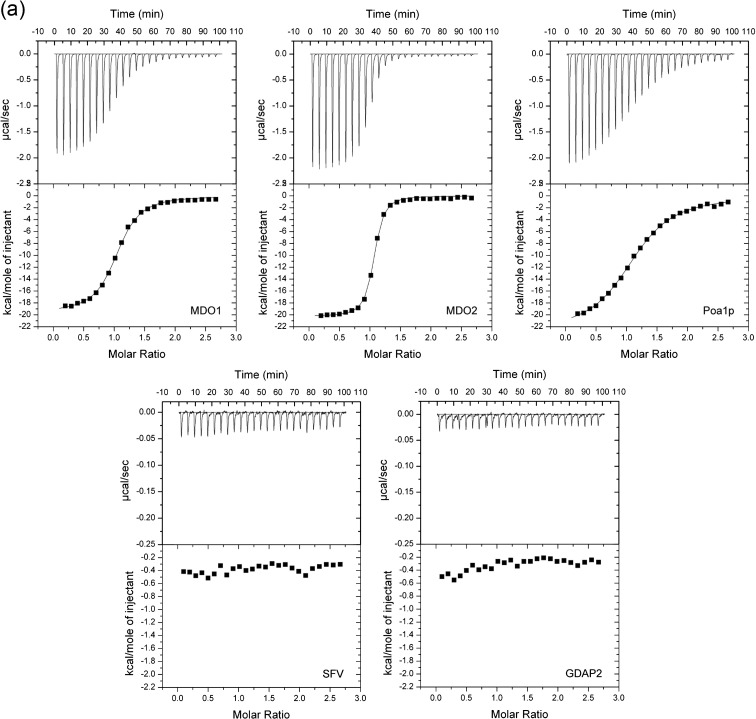
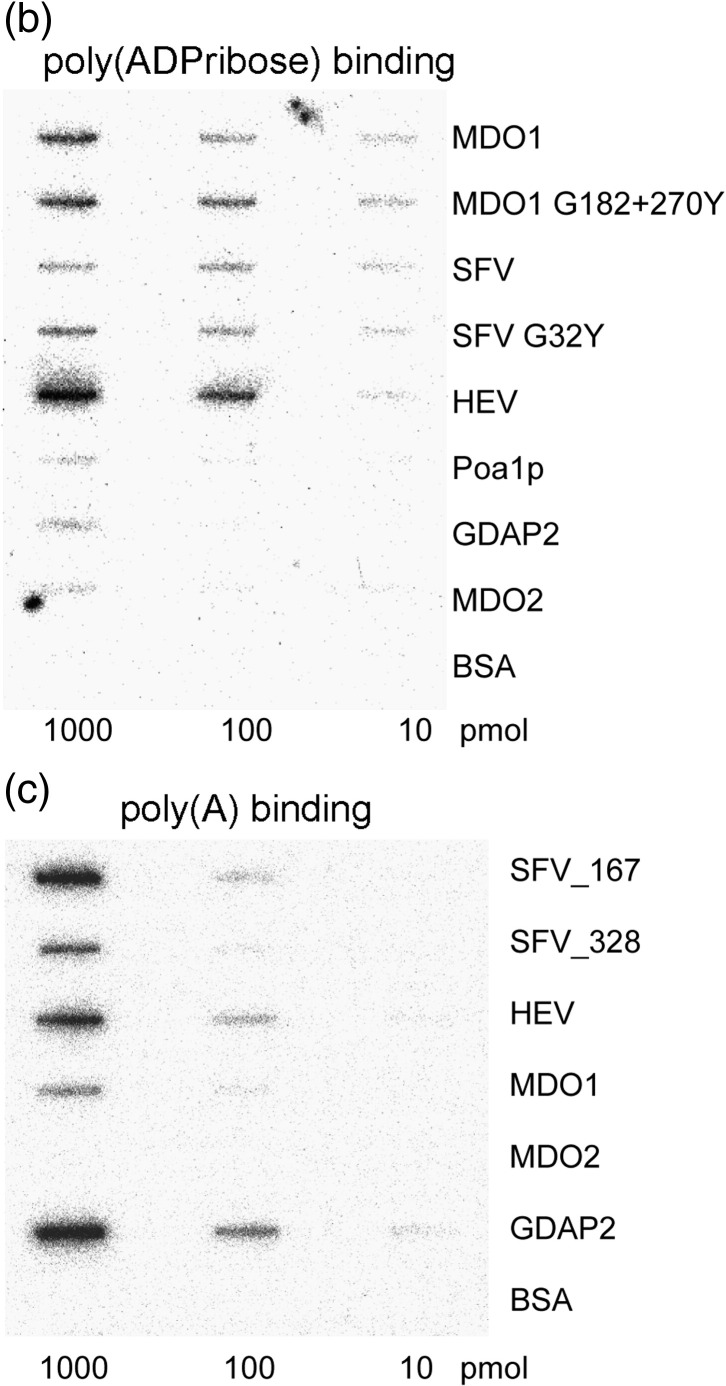
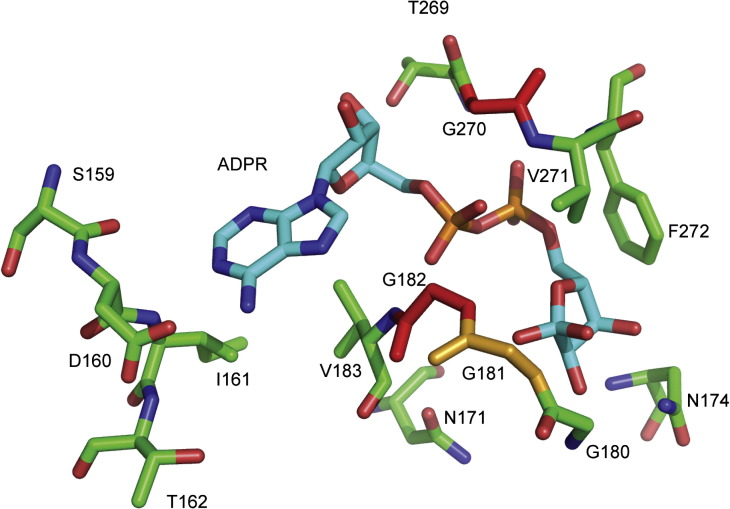
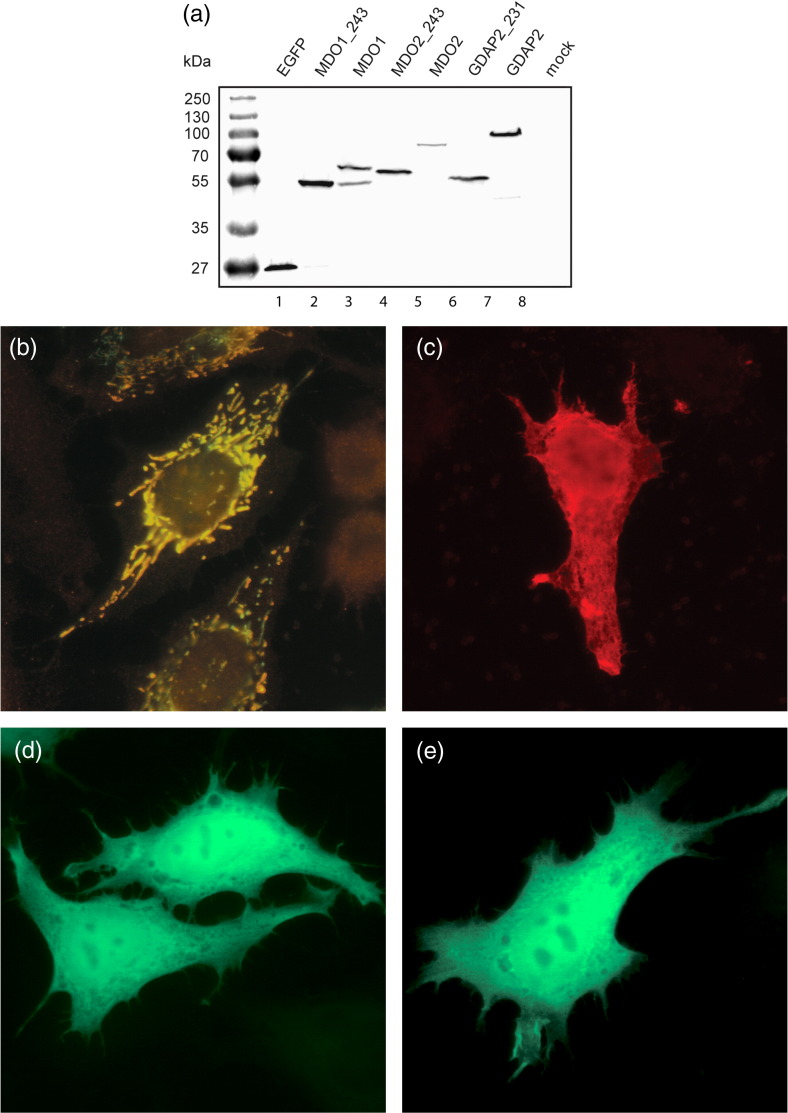
Macro domain is a highly conserved protein domain found in both eukaryotes and prokaryotes. Macro domains are also encoded by a set of positive-strand RNA viruses that replicate in the cytoplasm of animal cells, including coronaviruses and alphaviruses. The functions of the macro domain are poorly understood, but it has been suggested to be an ADP-ribose-binding module. We have here characterized three novel human macro domain proteins that were found to reside either in the cytoplasm and nucleus [macro domain protein 2 (MDO2) and ganglioside-induced differentiation-associated protein 2] or in mitochondria [macro domain protein 1 (MDO1)], and compared them with viral macro domains from Semliki Forest virus, hepatitis E virus, and severe acute respiratory syndrome coronavirus, and with a yeast macro protein, Poa1p. MDO2 specifically bound monomeric ADP-ribose with a high affinity (K(d)=0.15 microM), but did not bind poly(ADP-ribose) efficiently. MDO2 also hydrolyzed ADP-ribose-1'' phosphate, resembling Poa1p in all these properties. Ganglioside-induced differentiation-associated protein 2 did not show affinity for ADP-ribose or its derivatives, but instead bound poly(A). MDO1 was generally active in these reactions, including poly(A) binding. Individual point mutations in MDO1 abolished monomeric ADP-ribose binding, but not poly(ADP-ribose) binding; in poly(ADP-ribose) binding assays, the monomer did not compete against polymer binding. The viral macro proteins bound poly(ADP-ribose) and poly(A), but had a low affinity for monomeric ADP-ribose. Thus, the viral proteins do not closely resemble any of the human proteins in their biochemical functions. The differential activity profiles of the human proteins implicate them in different cellular pathways, some of which may involve RNA rather than ADP-ribose derivatives.
Figures
References
-
- Allen M.D., Buckle A.M., Cordell S.C., Löwe J., Bycroft M. The crystal structure of AF1521 a protein from Archaeoglobus fulgidus with homology to the non-histone domain of macroH2A. J. Mol. Biol. 2003;330:503–511. - PubMed
-
- Martzen M.R., McCraith S.M., Spinelli S.L., Torres F.M., Fields S., Grayhack E.J., Phizicky E.M. A biochemical genomics approach for identifying genes by the activity of their products. Science. 1999;286:1153–1155. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
